- Учителю
- Научная работа на тему Эволюция и происхождение птиц
Научная работа на тему Эволюция и происхождение птиц
ВВЕДЕНИЕ
Птицы как особый класс позвоночных животных сформировался примерно 80-100 млн. лет назад. Наиболее ранние ископаемые останки веерохвостых птиц имеют возраст около 120-135 млн. лет. В ходе эволюции у птиц сформировались сложные формы поведения и взаимосвязей с окружающей средой, что определило огромное многообразие форм представителей этого класса. Высокий уровень обмена веществ, разнообразие трофических связей и способность большинства видов к активному полету или плаванию позволили птицам за относительно короткий период освоить все территории и акватории Земли. Большое многообразие видов и высокая численность многих из них (десятки и сотни миллионов особей) определяют значительную роль птиц в биогеоценозах. Многие птицы приспособились к жизни в урбанизированных ландшафтах и в поселениях человека. Жизнедеятельность ряда видов птиц-синантропов стала одной из важных проблем, особенно в больших городах. С птицами связано распространение некоторых болезней человека и домашних животных. Многие виды птиц имеют большое значение как объекты охоты. В ряде районов развит регулярный сбор в пищу яиц или птенцов колониальных птиц.
Внимание международной общественности к вопросам охраны птиц в значительной степени объясняется не только изменением и оскудением авифауны в некоторых странах, но и особенностями сезонного распределения птиц. Для многих видов птиц места гнездования и районы зимовки удалены на сотни и даже тысячи километров. Во время регулярных сезонных миграций эти птицы оказываются на территории различных стран, поэтому сохранение или рациональное использование отдельных видов или групп видов птиц требует согласованных мер между различными странами. Изменения условий обитания вызывали неоднократные перемещения птиц. Так, длительные ледниковые периоды в северном полушарии определили относительную бедность современной авифауны Палеарктики и Неарктики. В настоящее время в соответствии с концепцией широкого биологического вида общее число видов птиц в фауне мира примерно 8600-9000. Они объединены в 27 отрядов. Из них 769 видов из 20 отрядов зарегистрировано в авифауне России. Уничтожение или изменение местообитаний таких видов может привести к резкому сокращению их численности и даже полному вымиранию. В то же время имеется много видов или хорошо обособленных подвидов птиц, которые по сравнению с другими родственными видами весьма малочисленны в наше время. Они могли быть редкими издавна или стали редкими в последние годы, преимущественно в результате антропогенного воздействия. Для таких птиц необходимо учитывать тенденции изменения численности и выяснять минимальный состав популяции - предел, ниже которого вид или другой таксон может оказаться на грани исчезновения. Важное, значение имеет тенденция изменения численности вида и его ареала. Одни виды увеличиваются в числе или расширяют ареал, другие остаются на стабильном уровне, численность третьих, как и их распространение, сокращается. И если сокращение численности и распространение вида не являются естественными циклическими изменениями, а наблюдается в течение ряда лет неуклонное уменьшение популяции, то такие виды заслуживают внимания как возможные кандидаты в исчезающие.
Актуальность темы: Эволюционные преобразования, приведшие к появлению класса - Aves, в последние десятилетия привлекают особое внимание палеонтологов. Недавние многочисленные находки значительно расширили знания о морфологии и диверсификации ранних птиц и их возможных предков, а обширный сравнительно-анатомический материал по рецентным видам представляет собой базу для реконструкции эволюционных преобразований, приведших к появлению современных групп птиц.
Цель работы: заключается в разработке и демонстрации системного морфо-функционального, морфо-экологического подхода к изучению и реконструкции эволюции и адаптивных преобразований аппарата двуногой локомоции птиц, а также уникальная способность птиц к полету. С этой целью поставлены следующие задачи:
-
создание исторического обзора по теме с выявлением современных направлений в исследованиях;
-
анализ наиболее значимых для реконструкции адаптивной эволюции задних конечностей птиц структур;
-
вскрытие ключевых адаптаций, экологических возможностей и адаптивной эволюции аппарата двуногой локомоции ряда рецентных групп птиц (Кукушкообразных, Ракшеобразных, Трогонообразных, Дятлообразных, Воробьинообразных);
-
экологические, физиологические и биохимические исследования функционирования систем, органов и тканей, активно участвующих в процессах адаптации к специфическим факторам больших и малых высот.
Происхождение и эволюция птиц
Глава 1. Исторический обзор.
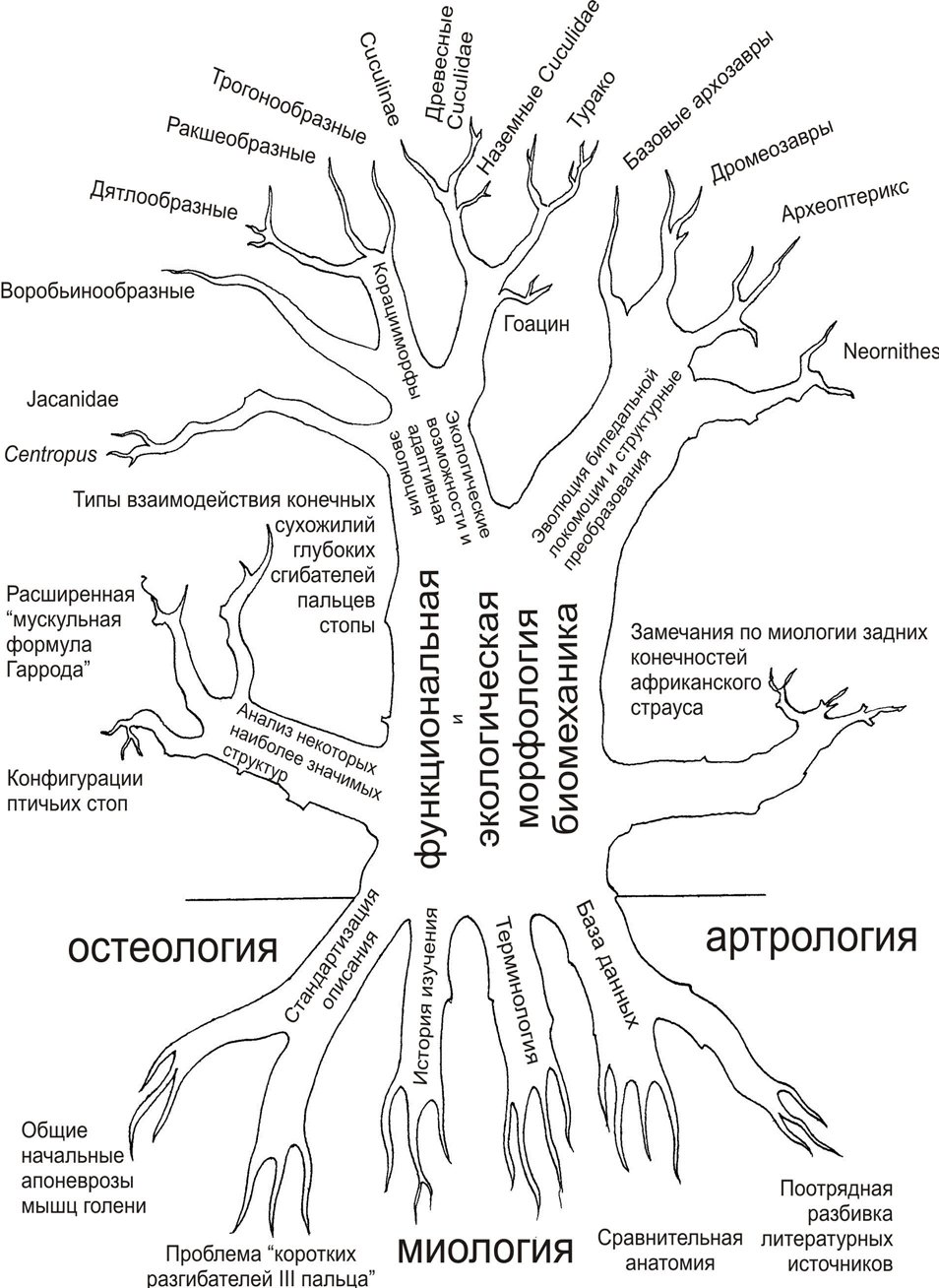
Одна из загадок, вызывающая среди ученых жаркие споры, -происхождение птиц. Обширная работа по изучению теорий и гипотез происхождения и эволюции птиц кристаллизуется в некое логическое целое, объясняемое причинно-следственной необходимостью интерпретации фактов, но и самой историей исследования. Они, подобно боковым корням или придаточным побегам растения, намечают и определяют направления дальнейшего роста исследования вширь (рис.1. Древо, иллюстрирующее структуру и логику исследований). [1].
Палеонтологические материалы по птицам очень скудны и отрывочны. Поэтому говорить о предках птиц среди рептилий и о происхождении и взаимоотношениях современных групп птиц приходится преимущественно предположительно, основываясь на морфологических особенностях современных групп и, в очень малой степени, на отрывочных палеонтологических данных.
По современным представлениям, предками птиц
были архозавры Аrchosauria - обширная и очень разнообразная группа
рептилий, господствовавшая в мезозое и включавшая различных
динозавров, крокодилов, летающих ящеров. Непосредственных предков
птиц, видимо, нужно искать среди наиболее древней и примитивной
группы архозавров - текодонтов или псевдозухий Thecodontia
(Pseudosuchia), давших начало и остальным более
высокоорганизованным группам архозавров. Псевдозухии по внешнему
виду были похожи на ящериц. Они вели преимущественно наземный образ
жизни, питались, видимо, разнообразной мелкой животной пищей.
Задние ноги были несколько длиннее передних, и при быстром беге
животные опирались о почву только задними конечностями. По многим
морфологическим особенностям псевдозухии весьма сходны с птицами
(детали строения черепа, таза, задних конечностей). Промежуточных
форм, которые позволяли бы выяснить стадии развития оперения и
выработку способности к полету, пока не найдено. Полагают, что
переход к древесному образу жизни и приспособление к прыжкам с
ветки на ветку сопровождался разрастанием и усложнением структуры
роговых чешуй на боках туловища и хвоста, по заднему краю передних
и задних конечностей, что обеспечивало возможность планирования и
удлинения прыжков (гипотетическая стадия предптицы, по Хейльманну,
1926, рис.2). [1].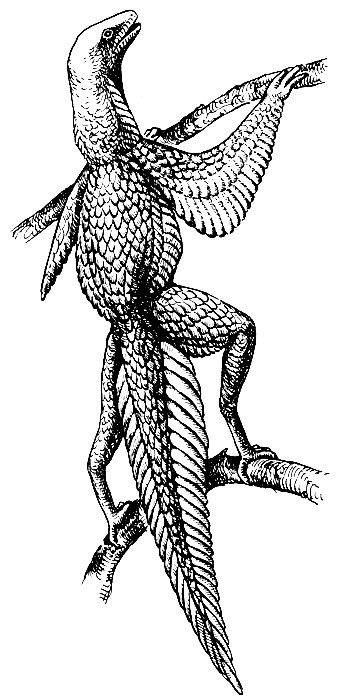
Рис. 2. Гипотетический предок птиц - предптица (по Хейльманну, 1926).
Дальнейшая специализация и отбор привели к развитию оперения крыла, обеспечившего возможность сначала планирующего, а затем активного полета. Оперение, покрывающее все тело птиц, вероятно, развивалось не столько как приспособление к полету (обеспечение обтекаемости тела), сколько как приспособление к термоизоляции; высказываются предположения, что оно могло сформироваться еще до выработки способности к полету. У некоторых псевдозухий были удлиненные роговые чешуи с четким продольным гребнем, от которого отходили мелкие поперечные ребрышки. Такие чешуйки, видимо, можно рассматривать как структуру, из которой путем расчленения могло возникнуть птичье перо. Обособление птиц от пресмыкающихся, видимо, произошло уже в конце триаса - начале юры (170 - 190 млн. лет назад). Однако ископаемых останков птиц этого времени пока не найдено. Довольно хорошие остатки двух самых древних известных нам птиц были найдены в 60-х годах XIX в. у Золенгофена (Бавария) на разработках сланцевых песчаников, представляющих отложения мелководного залива юрского моря (возраст около 150 млн. лет). Их назвали Archaeopteryx lithographica и Archaeornis simetisi. В 1956 г. там же (в радиусе до 300 ж, но на иной глубине) найден значительно худшей сохранности отпечаток еще одной особи. Часть исследователей рассматривает эти находки как остатки трех разных видов (два вида рода Archaeopteryx и один рода Archaeornis), другие же считают представителями одного вида Archaeopteryx lithographica, объясняя различия между особями как индивидуальные, возрастные и половые. Вопрос этот нуждается в дополнительном изучении, но и сейчас совершенно очевидна очень большая близость всех этих особей. Все они имеют хорошо развитое оперение крыльев, хвоста и тела, некоторые пропорции черепа ближе к птичьим, чем к рептильным, птичьи черты несут пояс передних конечностей и таз, задние конечности. Типично рептильные черты: нет рогового клюва, имеются слабые зубы, кисть несет первостепенные маховые, но пряжка еще не образовалась и есть три хорошо развитых подвижных пальца с крупными когтями, образование сложного крестца только началось (позвонки еще не срастаются, подвижные кости соединяются с небольшим числом позвонков), бедро причленяется под тупым углом (направлено несколько вбок), проксимальные тарзальные кости еще не приросли к концу большой берцовой кости, хорошо развита малая берцовая кость, не завершено образование цевки и т. п. По внешнему облику, ряду внутренних морфологических признаков и, насколько это можно предполагать сейчас, образу жизни архептерикс (и археорнис) - древние, примитивные, но птицы (рис.3 реконструкция внешнего облика древних птиц).



Рис.3. реконструкция внешнего облика древних птиц.
Подкласс настоящих, или веерохвостых, птиц Neornithes, видимо, - прямые потомки каких-то ящерохвостых птиц Archaeornithes. Некоторые исследователи (например, Гадов Г.Ф.) считают за прямых предков птиц археоптерикса и археорниса, другие (их, пожалуй, большинство) рассматривают группу археоптерикса как примитивную, но специализированную боковую и слепую ветвь, полагая, что предками современных птиц должны быть другие, пока еще не обнаруженные ящерохвостые птицы (в случае их обнаружения они, вероятно, образуют другой отряд этого подкласса).
Происхождение и взаимоотношения современных групп (отрядов) птиц остаются, как уже говорилось, за отсутствием достаточных палеонтологических материалов, как правило, очень гипотетическими. Наибольшее внимание уделил этому вопросу М. Фюрбрингер, заметно меньшее - Г. Гадов. Не внесли много нового и бесспорного в эту проблему и современные авторы, в той или иной степени касавшиеся филогении птиц. Поэтому здесь изложено представление о взаимоотношениях отдельных групп птиц преимущественно по Фюрбрингеру, но в ряде случаев, где это нужно, внесены некоторые изменения (в частных случаях эти изменения специально не оговариваются). Ископаемые остатки веерохвостых птиц обнаружены только в меловом периоде (их возраст примерно 80 - 90 млн. лет), т. е. они на несколько десятков миллионов лет моложе археоптериксов. По нескольким плохой сохранности остаткам описаны гесперорнисы - очень крупные (длиной более 100 см) птицы, внешне, видимо, несколько похожие на гагар или поганок, совершенно не летавшие (грудина без киля, от передней конечности сохранилось только маленькое тоненькое плечо), но, вероятно, они хорошо плавали и ныряли: челюсти их несли зубы. В меловых же отложениях найдены остатки других зубастых птиц - ихтиорнисов. Они имели хорошо развитый киль грудины, типичное птичье крыло и, видимо, обладали активным полетом. По размерам они были примерно с голубя. Родственные отношения зубастых меловых птиц с другими птицами очень неясны. Возможно, часть этих остатков относится к меловым динозаврам, а не птицам, но часть, несомненно, зубастые птицы. Фюрбрингер сближал гесперорнисов с гагарами и поганками, а ихтиорнисов - с чайками, но при этом подчеркивал их очень давнюю обособленность. Гадов Г.Ф. гесперорнисов выделял в надотряд, показывая этим их обособленное положение, а ихтиорнисы как самостоятельный отряд (тоже обособленный) начинали перечень отрядов килегрудных птиц. Подчеркивая неясность происхождения (да и объем) этих групп, Уэтмор А. (1960) выделяет их в самостоятельные надотряды. [2].
Из меловых отложений описано еще несколько семейств птиц (по обломкам челюстей и трубчатых костей), но их положение и внешний облик очень неясны: возможно, хотя бы часть этих находок - остатки рептилий. Встреченные в отложениях третичного периода кайнозойской эры (возраст менее 55 млн. лет) останки птиц с большей или меньшей вероятностью могут быть отнесены к современным отрядам. Во всяком случае, они принадлежат к типичным птицам, близким к тем или иным группам современных птиц. Судя по этим материалам, можно предполагать, что интенсивная адаптивная радиация птиц и становление большинства современных отрядов проходило в самом конце мелового периода мезозойской эры - в третичный период кайнозойской эры, т. е. в период примерно 70 - 40 млн. лет назад. [1].
Гадов Г.Ф. всех ныне живущих и ископаемых крупных нелетающих страусоподобных птиц выделял в надотряд бескилевых, подчеркивая этим их сходство и филогенетическое родство (т. е. монофилию), а также их определенную обособленность от остальных групп птиц. Среди килевых птиц наиболее близки к страусоподобным тинаму (скрытохвосты), куриные, журавлеобразные и близкие к ним группы. Фюрбрингер и большинство современных авторов (Уэтмор, Штреземанн и др.) считают страусоподобных птиц сборной полифилетической группой, разбиваемой на ряд самостоятельных отрядов, начинающих систему, что подразумевает их определенную примитивность и изолированность. Их общие сходные черты - результат конвергенции: крупные размеры, потеря способности к полету, приспособление к быстрому бегу и т. д.
По Гадову, эволюция остальных птиц шла в двух направлениях (две ветви, см. выше), каждое из которых в свою очередь разделяется на 2 группы родственных отрядов. Поганко-гагарообразные (ихтиорнисы, поганки, гагары, пингвины и трубконосые) родственны группе Pelargomorphae (аистообразные, гусеобразные, хищники), а близкая к страусоподобным группа курообразных (тинаму, куриные, журавлеобразные, куликообразные) дала начало ракшеобразным (кукушки, ракши, воробьиные). [3].
Эволюционные представления Фюрбрингера в общих чертах близки к этой схеме Гадова (о иных взглядах на страусоподобных говорилось выше), но он излагает свои представления с большей степенью детализации. Крупный ствол птиц - отряд Pelargornithes - делится на ряд ветвей: гусеобразных, поганкообразных (включая поганок, гагар, гесперорнисов); от основания этих ветвей отходит еще одна мощная ветвь, которая, разделяясь, дает начало веслоногим, дневным хищникам и группам подотряда аистообразных.
Отряд паламедей Palamedeiformes занимает промежуточное положение между нанду и отрядом Pelargornithes. Промежуточные отряды пингвинов Aptenodytiformes и трубконосых Procellariiformes довольно обособлены; возможно, они имеют давние слабые связи друг с другом и, еще более отдаленные, с отрядами Pelargornithes и куликообразных Charadriornithes. Ихтиорнисы Ichthyornithiformes - тоже промежуточный отряд, но с несколько более выраженными давними связями с куликообразными. Отряд куликообразных Charadriornithes очень давними связями видимо может быть приближен к отряду ракшеобразных. Основные ветвления ствола куликообразных - подотряд Laro-Limicolae - кулики, чайки и чистики; к ним довольно близки зобатые бегунки и дрофы (переходной группой между куликами и дрофами служат авдотки). От основания ствола куликообразных обособились еще две крупные ветви: журавлеобразные Gruiformes и пастушковые Ralliformes. Журавлеобразные разделяются на несколько ветвей: собственно журавлиных (журавли, арамиды, трубачи, кариамы) и занимающих несколько обособленное положение солнечных цапель и кагу. Пастушковые подразделяются на родственные ветви: пастушков, лапчатоногих и стоящих несколько обособленно трехперсток и мадагаскарских пастушков.
Один из древних стволов родословного древа птиц - отряд курообразных Alectorornithis. Наиболее примитивная ветвь - подотряд бескрылых, или киви Apterygiformes, включающий киви и моа; обе группы разошлись почти сразу после своего обособления. Общим происхождением с предыдущей ветвью, видимо, связаны тинаму (или скрытохвосты) Crypturiformes, в свою очередь через предков близкие к подотряду куриных Galliformes (практически в современном объеме). Отряд курообразных очень давними родственными связями, вероятно, более или менее близок к пастушковым Ralliformes, а через них и ко всему отряду куликообразных.
Промежуточные отряды голубеобразных Columbiformes (включающий рябков и голубей) и попугаев Psittaciformes занимают в системе обособленное положение. Вероятно, через очень давние отношения они как-то связаны с обширным отрядом ракшеобразных. По представлениям Гадова, голуби и рябки в ранге подотрядов входят в отряд куликообразных, а попугаи - тоже как подотряд - в отряд кукушкообразных. По сути дела сходной точки зрения придерживается и Уэтмор, ставящий отряд голубеобразных сразу же за отрядом куликообразных, а отряд попугаев - рядом с отрядом кукушек. [3].
Обширный и разнообразный отряд Coracornithes своими истоками связан с предками отряда куликообразных. Основная и наиболее мощная ветвь - подотряд дятлов и воробьиных Pico - Passeriformes, которая затем разделяется на отдельные группы собственно воробьиных и дятловых. От основания этой ветви обособились Macrochires (стрижи, колибри) и птицы мыши Colii. Еще раньше от этой ветви обособляются трогоны Тгоgones. От самого основания ветви дятлово-воробьиных птиц обособляются и три других подотряда ракшеобразных. Подотряд кукушкообразных Coccygiformes отделяет ветвь якамар Galbulae (якамары и пуховки, а по Уэтмору -это подотряд отряда дятлов), а затем разделяется на кукушек Cuculidae и бананоедов Musophagidae. Подотряд ракшеобразных Со- raciiformes разделяется на собственно ракш Coraciae и на вторую ветвь, вскоре делящуюся на сов Striges и козодоев Caprimulgi. Подотряд зимород- ковых Halcyoniformes (он, видимо, ближе к Pico-Passeriformes, чем к другим подотрядам), отделяя в самом начале боковую веточку тоди Todi (кроме тоди Фюрбрингер сюда включает и момотов), разделяется на 3 родственные группы: зимородковых Halcyones (Alcedinidae), удодов Bucerotes (удоды, птицы-носороги) и щурок Meropes. [1].
Сопоставление филогенетических представлений Фюрбрингера со взглядами Гадова и Уэтмора (очередность и порядок перечисления отрядов) показывает значительное их совпадение; резкие отличия в положении и взаимоотношении отдельных групп указаны выше.
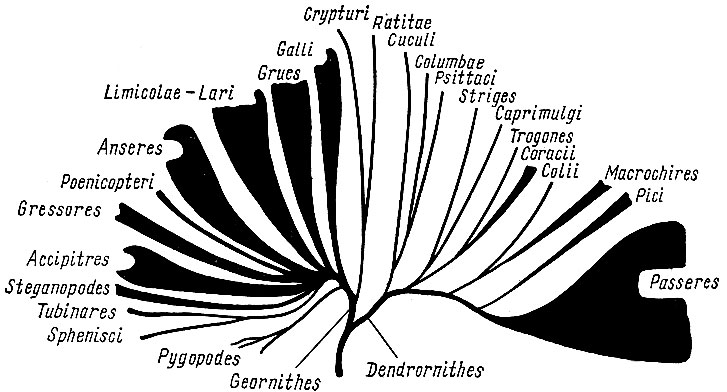
Рис. 4. Схема филогенетических отношений в классе птиц (по Берндту
и Майзе, 1960)
В последние годы попытку графического изображения филогенетических взаимоотношений между отдельными группами птиц предприняли немецкие орнитологи Берндт и Майзе в капитальной трехтомной сводке "Естественная история птиц" (Bernd R.,Meise W. Naturgeschichte derVogel, Kosmos, Stuttgart, 1960 (1962), v. 2, стр 668 - 673). Подкласс древних ящерохвостых птиц Palaeornithes дал начало подклассу новых веерохвостых птиц Neornithes, который включает 24 отряда, подразделяемых авторами обычно сразу на семейства; таксономические категории ранга подотряда и надсемейства используются, как правило, только в отряде воробьиных. Взаимоотношения между отрядами птиц, по Берндту и Майзе, выглядят следующим образом: от древних птиц эволюция шла по двум направлениям (ветвям) - ветви (надотряд) наземных и водных птиц Geornithes и ветви (надотряд) древесных птиц Dendrornithes. От самого начала ветви наземных и водных птиц (рис. 4.) общим стволом отделились, позже разделившись, тинаму Crypturi и бескилевые птицы Ratitae, включая всех ныне живущих страусоподобных, в том числе и киви, и ископаемых моа и эпиорнисов. Далее обособляются куриные птицы Galli (в полном объеме отряда Уэтмора) и на этом же уровне, но самостоятельной ветвью - гагарообразные Pygopodes (включающие ископаемых меловых зубастых птиц гесперорнисов, гагар и поганок). Все остальные отряды этой ветви (надотряда) имеют общее происхождение - представляют собой как бы мутовку стволов, отходящих от одного участка. Относительно тесно связаны друг с другом и несколько противопоставляются другим группам отряды журавлеобразных (пастушки, мада- гаскарские пастушки, солнечные цапли, лапчатоноги, кагу, настоящие журавли, арамы, трубачи, дрофы, кариамы, трехперстки; из ископаемых - диатримы и форораки Phororhacidae) и болотно- и чайкообразных птиц Limicolae - Lari (яканы, различные группы собственно куликов, чайки чистики и - из ископаемых - зубастые меловые птицы - ихтиорнисы). Далее от общего основания мутовки (т. е. имея общих предков) последовательно обособляются отряды гусеобразных Anseres (паламедеи и собственно гусеобразные), фламинго Phoenicopteri, голенастые Gressores, дневные хищники Accipitres и веслоногие Steganopodes. От форм, близких к предкам веслоногих, отделяется ветвь, которая затем разделяется на отряды трубконосых Tubinares и пингвинов Sphenisci. [2].
Развитие второй ветви (надотряда) древесных птиц Dendrornithes начинается от каких-то предковых форм, видимо, более древних, чем предки первой ветви (первого надотряда). Сначала общим стволом обособляются 3 отряда: кукушкообразных Cuculi, голубей Columbae (голубки, рябки и дронты) и попугаев Psittaci. Следующая обособившаяся ветвь разделяется на 2 ответвления, каждое из которых образует по 2 отряда. Одно из них дает начало совам Striges и козодоям Caprimulgi, другое - трогонам Trogones и ракшеобразным Coracii. Дальше от общего ствола также общей ветвью обособляются отряды птиц-мышей Colii и стрижеобразных Macrochires (стрижи и колибри), самостоятельной ветвью отходит отряд дятлов Pici (в объеме Уэтмора), а основной ствол ветвится на многочисленные подразделения отряда воробьиных Passeres. Помимо этого, авторы дают гипотетические схемы, в которых пытаются представить филогенетические взаимоотношения в пределах нескольких отдельных отрядов (дневных хищников, веслоногих, гагарообразных и воробьиных).
Филогенетические схемы Берндта и Майзе ясны, наглядны и легко запоминаются. Однако они практически совсем не аргументированы, не обоснованы материалами и, видимо, в значительно меньшей степени отражают действительные родственные (филогенетические) отношения между отдельными группами птиц, чем соответствующие схемы Фюрбрингера. Теоретически маловероятно, что уже на начальных стадиях эволюции класса четко определилась последующая биотопическая специализация птиц, что выражается в изложенной схеме деления подкласса "новых птиц" только на 2 обособленных надотряда и в почти линейном, последовательном обособлении отрядов. Оценивая по современным формам присущий птицам характер эволюции, выражающийся в широкой адаптивной радиации в пределах отдельных групп (о чем кратко упоминалось выше), кажется более логичным предполагать, что подобная же широкая адаптивная радиация была характерна и для начальных фаз эволюции класса. Если это так, то можно говорить, что филогенетические схемы Фюрбрингера хотя бы внешне действительно отражают преобладание такого типа эволюции.
Сейчас еще рано говорить о вполне сложившейся системе класса. Однако системы Фюрбрингера, Гадова и Уэтмора, видимо, могут рассматриваться как хорошая основа для построения естественной системы. Их материалы дают и самое общее представление о филогенетических взаимоотношениях большинства групп. [3].
Глава 2. Анатомическое и морфологическое описание.
В начале XXI века продолжаются исследования происхождения и эволюция птиц, которые можно условно разделить на несколько направлений:
Первое объединяет работы, привлекающие особенности морфологии задних конечностей (как правило, наряду с другими категориями признаков) для выяснения филогенетических, а точнее, кладистических отношений между таксонами, как вымерших, так и современных птиц (Hughes, 2000; Cracraft, Clarke, 2001; Chiappe, Dyke, 2002; Mayr, 2002, 2003; Mayr, Clarke, 2003; Dyke et al., 2003; Ericson et al., 2003; Zhou, 2004; Moyle, 2005; Manegold, 2005 и др.).
Второе касается изучения биомеханики двуногой локомоции птиц. В этом направлении следует отметить работы Аник Абурашид с коллегами, в которых исследование кинематики сочетается с функциональным анализом некоторых особенностей строения скелета конечностей и, в итоге, служит основой для проектирования механических моделей-роботов (Abourachid, 2000, 2001a; Abourachid, Renous, 2000, 2001; Abourachid et al., 2001, 2004, 2006; Hugel et al., 2003).
Третье направление - ветеринарно-анатомическое. Основным объектом служат хозяйственно значимые виды птиц; исследования сконцентрированы на анатомии задних конечностей и основных локомоторных актах (Harcourt-Brown, 2000; Salzer, Starck, 2000; Williams et al., 2000; Buchanan, Marsh, 2001; Frey et al., 2001; Rinck, 2002; Linn et al., 2003; Ganglet al., 2004).
Четвертое направление - морфо-функционально-экологическое. Морфология локомоторного аппарата птиц здесь изучается в связи с его функционированием в рамках конкретных адаптаций. Традиционно-развиваемой русской морфологической школой, в последние годы оно нашло отражение в работах Богдановича, Дзержинского, Волкова, Зеленкова, Зиновьева, Ильинского (Богданович, 2000, 2007; Волков, 2006; Зеленков, 2006, 2007; Дзержинский, 2000; Ильинский). Эволюция отдельных групп птиц предстает в них в виде цепи конкретных адаптаций.
Когда птицы приобрели способность к полету, их внутреннее строение сильно изменилось по сравнению с предковым, свойственным рептилиям. Для уменьшения веса животного одни органы стали более компактными, другие были утрачены, а чешуи сменились перьями. Более тяжелые, жизненно необходимые структуры переместились ближе к центру тела ради улучшения его балансировки. Вдобавок повысились эффективность, скорость и регулируемость всех физиологических процессов, что обеспечило требуемую для полета мощность.
Общие черты строения рептилий и птиц:
-
нет кожных желез (кроме специализированных, например, копчиковой);
-
сильно развиты роговые кожные производные (перья, чешуя, роговой клюв, когти);
-
один затылочный мыщелок;
-
клиновидная и нижнечелюстная кости состоят из нескольких самостоятельных частей;
-
типичная нижняя скуловая дуга;
-
квадратная кость подвижна и имеет механизм передачи ее движений на надклювье;
-
развиты височная и предглазничная ямки;
-
длинный подвижный шейный отдел;
-
сложный крестец;
-
на ребрах имеются крючковидные отростки;
-
мезотарзальное сочленение на тазовой конечности;
-
сходное строение почки, печени и других органов.
Отличия современных птиц и рептилий:
-
лучше развит головной мозг (особенно полушария);
-
лучше развиты органы зрения и слуха;
-
теплокровные;
-
полное разделение артериальной и венозной крови;
-
наличие воздухоносных мешков, обеспечивает наилучшую вентиляцию легких;
-
большая подвижность;
-
наличие перьевого покрова способствует поддержанию постоянной температуры тела, а так же обтекаемости тела при полете;
-
кладка крупных яиц и забота о потомстве приводит к относительно большому выживанию особей.
2.1. Адаптивные (приспособительные) изменения скелета.
Кости птиц прочные и легкие, часто
пневматизированы, голова
облегчена благодаря отсутствию зубов. Шейный отдел позвоночника
удлинен и чрезвычайно подвижен, выполняя вместе с головой роль
переднего руля, хватательной конечности, и обеспечивая круговой
обзор крупным, но малоподвижным глазам. Грудопоясничный отдел
укорочен, малоподвижен, хвостовой отдел видоизменен, превращен в
основу для рулевых перьев. Грудная конечность преобразовалась в
крыло, что привело к значительным изменениям в скелете, особенно в
дистальных звеньях.
2.2. Адаптивные (приспособительные) изменения мускулатуры.
Мускулатура птиц расположена на теле неравномерно и
обеспечивает
две основные функции - полет и хождение. Грудная конечность
приспособилась к движению в воздушной среде, благодаря чему только
мышцы крыла получили значительное развитие, остальная же
мускулатура в своем развитии сведена до минимума. [3]
2.3. Адаптивные изменения внутренних органов.
Внутренние органы располагаются таким образом, что наиболее
массивные из них лежат вблизи центра тяжести тела (печень,
мышечный
желудок). Облегчение системы органов пищеварения достигается
редукцией
некоторых ее участков (зубов, ободочной кишки), укорочением
кишечника и
усилением процессов всасывания (наличие ворсинок в толстом
кишечнике).
Система воздухоносных мешков облегчает массу тела, пневматизирует
кости и полость тела, создает условия для дополнительной аэрации. В
мочевыделительной системе отсутствует мочевой пузырь.
Концентрированная моча выводится вместе с фекалиями. Кладка яиц
и
внешнее развитие зародыша приводит к тому, что самки не
утяжеляются
плодом.[3].
2.4. Деление тела птицы на отделы.
Тело птицы можно разделить на осевую часть и конечности. В
осевой
части различают голову, шею, туловище и хвост. Каждая из указанных
частей делится на ряд областей (рис.5).
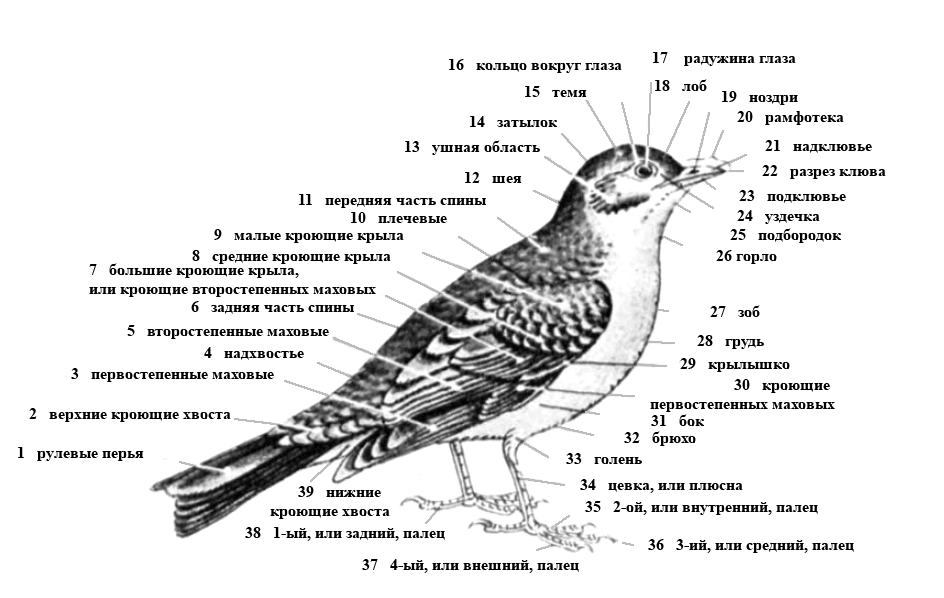
Рис. 5. Топография частей тела птицы.
Голова делится на мозговой и лицевой отделы. В
мозговом отделе различают области: лобную, теменную, затылочную,
надглазничную или височную, уха или околоушную, с ушной мочкой и
глаза. В лицевом отделе различают следующие области: щечную (щеки)
или уздечку - пространство между глазом и основанием клюва,
подбородочную с сережками у кур и индеек, клюва, состоящего из
верхней и нижней створок. Место соединения створок межу собой
называется разрезом клюва, а углы основания клюва - углом рта. На
верхней створке клюва проходит хребет или конек клюва, хорошо
выраженный у куриных. Костную основу области головы составляют
кости черепа. Шея делится на верхнюю шейную область или горло,
нижнюю шейную область или зоб, боковую шейную и заднюю шейную
области или зашеек. Костная основа всех областей шеи - шейные
позвонки. Туловище состоит из грудного, брюшного, тазового отделов.
Грудной отдел включает в себя области: переднюю грудную или хлуп,
костная основа которой - грудная кость и стернальные части ребер;
боковую с костной основой в виде ребер; переднюю спинную с костной
основой в виде грудных позвонков.
Брюшной отдел состоит из брюшной области или живота, не имеющей
костной основы. Задняя часть области живота, покрытая только
пуховыми перьями, называется кочнем. Тазовый отдел включает заднюю
спинную область, которая в основе своей имеет поясничные,
крестцовые и хвостовые позвонки, сросшиеся с подвздошной костью,
надхвостье и подхвостье, в основе которого лежат несросшиеся
хвостовые позвонки, а так же - хвост, костной основой которого
являются свободные хвостовые позвонки и пигостиль -
видоизменившиеся и сросшиеся последние хвостовые позвонки, к
которым прикрепляются рулевые перья хвоста. Грудная (передняя) и
тазовая (задняя) конечности птиц делятся на пояса и свободные
конечности. В свободных конечностях различают стило-, зейго- и
автоподий. Костной основой области плечевого пояса является
лопатка, коракоидная кость и ключица. Стилоподий грудной конечности
представлен областью плеча. Костная основа его - плечевая кость.
Зейгоподий - область предплечья. Костная основа - лучевая и
локтевая кости. Автоподий - кисть. Состоит из области запястья,
костную основу которого составляют две кости запястья; области
пясти, костная основа - пястно-запястная кость, и пальцев. Пальцы у
птиц на грудной конечности сильно редуцированы. Сохранились второй,
третий и четвертый пальцы. [4].
В тазовой конечности область тазового пояса
имеет своей основой подвздошную, лонную и седалищную кости,
сросшиеся в одну безымянную кость. Эта область, являясь поясом
тазовой конечности, входит как составная часть в тазовый отдел
осевого скелета в связи с прочным прикреплением подвздошной кости к
позвонкам. Свободная тазовая конечность состоит из тех же отделов,
что и грудная. Стилоподий тазовой конечности представлен
областью бедра, основа ее - бедренная кость; зейгоподий - область
голени, основа ее - большеберцово - заплюсневая и малоберцовая
кости; автоподий - стопой. В стопе различают область плюсны, или
цевку, и пальцы. Костную основу цевки (плюснозаплюсны) составляют
плюснозаплюсневая и первая плюсневая кости. Птицы обладают
двусторонней симметрией, что обусловливает определенное положение
частей тела и внутренних органов. Для определения расположения
части тела или органа в организме принято различать следующие
плоскости и направления:
1) Сагиттальная плоскость plani sagitalia - плоскость проведенная вертикально вдоль средины тела птицы от кончика клюва до конца пигостиля. Направление от любой сагиттальной плоскости наружу обозначается как латеральное (lateralis), а внутрь в сторону срединной плоскости - медиальное (medialis).
2) Фронтальные (дорсальные) плоскости plani
dorsalia - эти плоскости так
же проводятся вдоль тела птицы, но перпендикулярно сагиттальной,
т.е параллельно горизонтальной плоскости. По отношению к этой
плоскости рассматривают два направления: дорсальное (спинное) -
dorsalis - направление в сторону контура спины, и вентральное
(брюшное) ventralis - ориентированно в сторону живота.
3) Сегментальные (поперечные) плоскости plani
transversalia - эти плоскости проходят поперек тела птицы,
перпендикулярно продольным плоскостям, рассекая его на отдельные
участки (сегменты). По отношению к этим плоскостям рассматривают
два направления: к голове - краниальное cranialis, и к хвосту -
каудальное caudalis. На голове в этой плоскости различают оральное
oralis и аборальное направления aboralis.
Положение какой-либо части тела ближе к осевому скелету
относительно
другой части называется проксимальным, положение дальше от
осевого
скелета - дистальным. Передняя поверхность конечности
называется
спинковой - дорсальной, задняя (тыльная) сторона на грудной
конечности
(крыле) - пальмарной (волярной), на тазовой - плантарной. [4].
2.5. Скелет.
Скелет птиц подразделен на осевой (стволовой) и периферический (скелет конечностей). Осевой скелет включает в себя следующие отделы: лицевой и мозговой черепа, шейный, грудной, пояснично-крестцовый, хвостовой (рис. 6).
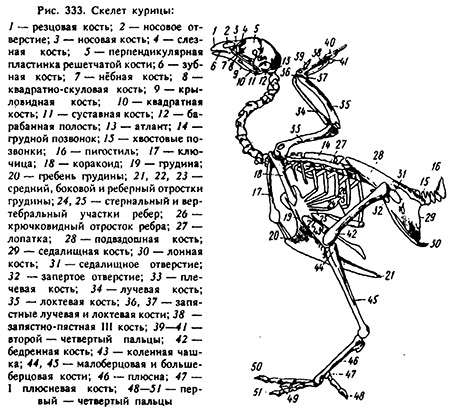
Рис.6. Скелет: 1. - резцовая (межчелюстная)
кость; 2 - ноздря; 3 - носовая; 4 - слезная; 5 - решетчатая; 6 -
зубная; 7 - квадратная кость; 8 - барабанная полость; 9 - атлант;
10 - коракоидная кость; 11 - ключица; 12 - лопатка; 13 - спинная
кость; 14 - астернальное и 15 - стрернальное ребра; 16 -
крючковидный отросток; 17 - тело; 18 - реберный, 19 - боковой и 20
- средний отростки грудины; 21 - ее гребень; 22 - плечевая, 23 -
лучевая, 24 - локтевая, 25 - запястно-локтевая и 26 -
запястно-лучевая кости, 27 - II палец; 28 -
пястные кости; 29 - III палец; 30 - IV палец; 31 - подвздошная
кость; 32 - седалищное
отверстие; 33 - хвостовые позвонки; 34 - пигостиль; 35 седалищная и
36 - лонная кости;
37 - запертое отверстие; 38 - бедренная кость; 39 - коленная чашка;
40 - малоберцовая; 41
- большеберцовая; 42 - плюсно-заплюсневая и 43 - I плюсневая кости;
44 - I палец; 45 - II
палец; 46 - III палец; 47 - IV палец.
Наибольшие особенности наблюдаются в строении позвоночного столба и черепа; также изменилось строение самой кости как органа. Облегченность скелета птиц создается за счет большей минерализации компакты, пористости губчатого вещества, пневматизации и раннего сращения костей. У самок перед яйцекладкой в костномозговых полостях трубчатых костей накапливается губчатая медуллярная кость - substantia spongiosa nova, которая при достаточном содержании Ca+2 в рационе заполняет всю полость кости. В процессе яйцекладки медуллярная кость расходуется на образование скорлупы. При недостатке Ca+2 истончается компакта, и кости становятся ломкими. [5].
Мозговой отдел черепа образован непарными затылочной, клиновидной, решетчатой и парными височной, теменной, лобной костями. Мозговой отдел не имеет межтеменной кости. Решетчатая кость не имеет лабиринта. Швы между костями черепа видны лишь в первые дни после вылупления. Под давлением крупных глаз глазничные крылья клиновидной кости срастаются между собой, с перпендикулярной пластинкой решетчатой кости и становятся межглазничной перегородкой. В результате мозговой отдел черепа не заходит рострально дальше глазниц. Затылочная кость имеет один мыщелок, что значительно увеличивает подвижность головы.
Лицевой отдел черепа образован парными резцовыми (межчелюстными), верхнечелюстной, носовой, слезной, небной, скуловой, крыловидной, квадратной, нижнечелюстной, непарным сошником и подъязычной костями.
Резцовые, верхнечелюстные и носовые кости образуют костный остов верхнего клюва - надклювье. Носовые кости имеют вид тонкой пружинистой пластинки, которая присоединяется (у гусиных суставом) к лобным и слезным костям и позволяет поднимать вверх надклювье. Движение это совершается одновременно с опусканием нижней челюсти - подклювья благодаря развитию нижней скуловой дуги и подвижности квадратной кости. Эта кость неправильной четырехугольной формы образует 4 сустава: с височной, крыловидной, скуловой и нижнечелюстной костями. Подвижное соединение крыловидной, скуловой, небной, квадратной, нижнечелюстной костей образуют хороший хватательный механизм птичьего клюва. В состав позвоночного столба - входят позвонки, которые наряду с наличием общих морфологических признаков имеют специфичные отличия, в том числе и видовые, в зависимости от их местоположения.
Шейный отдел. Позвонки подвижны, имеют короткие остистые и хорошо развитые поперечные отростки, рудименты ребер в виде реберных отростков. Сложный рельеф головок и ямок позвонков обеспечивает не только сгибание и разгибание, но и отведение в стороны, и ограниченное вращение. Ямка атланта сочленяется с одним суставным мыщелком затылочной кости. В атлантно-затылочном суставе возможны вращательные движения головы.
Грудной отдел. Первый и второй грудные позвонки соединяются между собой седловидным суставом, а со второго по пятый - срослись в сплошную кость, шестой грудной позвонок соединен подвижно как с пятым, так и с седьмым позвонком, в то время как седьмой грудной позвонок сросся с первым поясничным. У птиц первое, второе, реже третье ребро, а иногда и последнее ребра астернальные (не соприкасаются с грудиной), остальные - стернальные (соприкасаются с грудиной). От заднего края каждого ребра каудодорсально отходит крючковидный отросток, соединяющийся со следующим за ним ребром. Грудная кость сильно развита, пластинчатая и у кур имеет значительную вырезку по каудальному краю, у уток эта вырезка менее глубокая, а у гусей замыкается в отверстие. С вентральной стороны грудины у домашних птиц расположен гребень (киль) грудной кости. Грудная клетка у птиц конусовидна, значительно расширяется каудально. Реберных хрящей нет. Благодаря крючковидным отросткам ребра у птиц соединены между собой, более прочно, чем у млекопитающих.
Пояснично-крестцовый отдел. Поясничный и крестцовый отделы сливаются в один пояснично-крестцовый отдел, где 11-14 позвонков срастаются в пояснично-крестцовую кость - первый поясничный позвонок срастается с 7-м грудным, а задний крестцовый - с тремя - семью хвостовыми позвонками. С этими позвонками с двух сторон срастаются подвздошные кости. Отчего весь отдел называется тазовым.
Хвостовой отдел. Первые 5 позвонков соединены подвижно, а последние 4-6 срастаются, образуя копчик- плоскую треугольную косточку, к которой прикрепляются рулевые перья.
Скелет тазовой конечности: В тазовом поясе кости
таза сращены с пояснично-крестцовым отделом позвоночника. Лонные и
седалищные кости не срастаются между собой. Скелет свободной
конечности состоит из бедра, костей голени и стопы. Бедренная кость
пневматизирована. Из костей голени лучше всего развита
большеберцовая кость, она срастается с костями заплюсны, образуя
большеберцово-заплюсневую кость, которую иногда называют - беговой.
Беговая кость является самой мощной и длинной костью.
Кости стопы, кроме пальцев, срослись. Заплюсны не существует, так
как проксимальный ее ряд срастается с большеберцовой костью, а
центральная кость и дистальный ряд полностью сливаются с костями
плюсны. В плюсне срослись вторая, третья, четвёртая плюсневые
кости, которые вместе с костями заплюсны образовали
заплюснево-плюсневую кость - цевку.
У птиц обычно 4 пальца:
-
1-й палец задний висячий, состоит из 2-х фаланг,
-
2-ой палец - три фаланги,
-
3-й - четыре фаланги,
-
4-ый - пять фаланг.
Число пальцев и фаланг у различных птиц
неодинаково. У петухов на
плантарной поверхности цевки имеется шпорный отросток. [6].
2.6. Мышцы.
Скелетная мускулатура у птиц расположена на теле неравномерно (рис.
7).
Подкожные мышцы. Хорошо развиты, собирают кожу в
складки, что позволяет взъерошивать, поднимать и поворачивать
контурные перья.
Мышцы головы. Лицевая (мимическая) мускулатура отсутствует, жевательная мускулатура более дифференцирована, чем у млекопитающих, и хорошо развита. Имеются особые мышцы, действующие на квадратную кость, и другие подвижные кости черепа.
Мышцы стволовой части тела. Хорошо развиты в области шеи и хвоста. На шее много коротких и длинных мышц, расположенных в несколько пластов. Особенность позвонков - подвижность и большая длина шеи способствуют разгибанию, отведению и некоторому вращению не только шеи целиком, но и отдельных ее участков, в результате чего шея птицы принимает S-образный вид. Мышцы грудного и пояснично-крестцового отделов позвоночника не развиты в связи с их неподвижностью. Мышцы грудной клетки и брюшной стенки те же, что и у млекопитающих, кроме диафрагмы, которая имеет вид соединительнотканной пленки, не полностью отделяющей легкие от остальных органов. [2].
Рис.7. Мускулатура птицы.
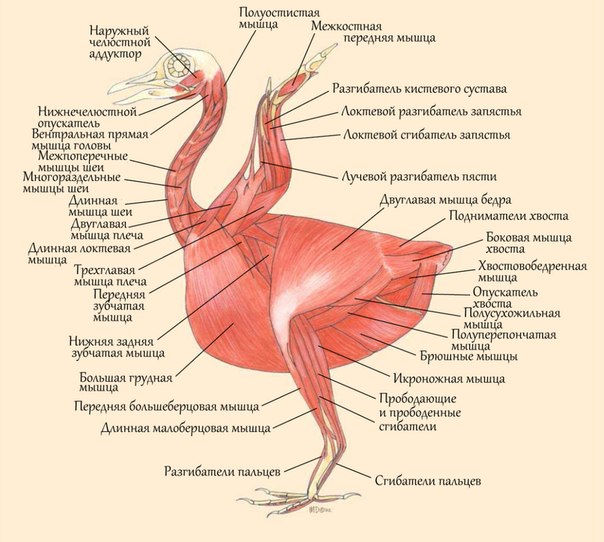
Мышцы грудной конечности. Сильно развиты и дифференцированы. В их число входит несколько десятков мышц. Грудная конечность птиц связана с туловищем не только суставами, но и с помощью мышц в области плечевого пояса и плеча. Это самые мощные мышцы тела. Они составляют до 45% от массы мускулатуры и выполняют основную работу во время полета, поднимая, опуская, супинируя (обеспечивая вращение передней (дорсальной) поверхности конечности наружу), пронируя (обеспечивая вращение передней (дорсальной) поверхности конечности внутрь) крыло в зависимости от маневра, совершаемого птицей (большая грудная мышца, подлопаточная, коракоидно-плечевая и др.)
Мышцы тазовой конечности. В области таза и бедра расположены разнообразные по функции мышцы, действующие на тазобедренный сустав. Из мышц, действующих на дистальные звенья конечности, развиты разгибатели и сгибатели. Их сухожилия обычно окостеневают. При движении благодаря сочетанному действию мышц на 2-3 сустава происходит одновременное разгибание и сгибание суставов. Сгибание всегда сопровождается приведением пальцев, разгибание - отведением. Этот механизм связывает суставы так, что при сгибании коленного сустава сгибаются и пальцы. [3].
2.7. Система органов кожного покрова.
Кожа покрывает все тело птицы, является барьером между организмом птицы и внешней средой. С одной стороны защищает организм от внешних воздействий, а с другой - осуществляет связь с внешней средой. В качестве
защитных приспособлений у птиц развиваются такие кожные производные, как перья и чешуйки. Через неповрежденную кожу в организм не проникают микробы и большинство вредных веществ. Связь с внешней средой осуществляется с помощью разнообразных чувствительных нервных окончаний, расположенных в коже, в результате чего она является обширным рецепторным полем. Кроме того, кожа депонирует до 30% крови и участвует в терморегуляции. При этом, через неё теряется 70-80% тепла, образуемого организмом птицы. Кожа птиц тонкая, сухая (из-за отсутствия потовых и сальных желез), имеет хорошо развитую подкожную клетчатку и образует складки. Железы представлены копчиковой железой, а у куриных еще и железами наружного слухового прохода. Кожа состоит из эпидермиса, собственно кожи или дермы и подкожного слоя. В разных участках тела строение кожи и толщина различны. Наиболее тонкая кожа - на оперенных участках (птерилиях), но и там она не одинакова. Наиболее толстая кожа на аптериях - участках, где кроющие перья не растут. На спине у сухопутных птиц кожа толще, чем на животе, а у водоплавающих - наоборот. Толстая кожа на подошве и в области межпальцевой перепонки. [5].
2.7.1. Роговые образования эпидермиса.
Чешуйки. У большинства птиц на тазовых конечностях от заплюсневого сустава до вершин пальцев перьевой покров не развит. У куриных эта область покрыта роговыми чешуйками, образующимися из сильно утолщенного слоя эпидермиса, содержащего твердый кератин. Роговой слой эпидермиса вдается гребнями в глубь кожи, отделяя чешуйки друг от друга. В гребнях кератин мягкий. На передней поверхности ноги чешуйки наиболее крупные и прикрывают друг друга, как черепицы. Самые мелкие чешуйки на медиальной и нижней частях плантарной поверхности ноги. У всех птиц в сетчатом слое, волокна образуют более тесные переплетения, чем в остальных участках кожи. У водоплавающих в области цевки и пальцев нет чешуек. Кожа в этих участках зернистая, напоминает шагреневую. Клетки рогового слоя содержат мягкий кератин.
Когти. Последняя фаланга каждого пальца снабжена роговым чехлом - когтем. Когти хорошо развиты у куриных и слабо - у гусиных. Роговой чехол образован толстым роговым слоем эпидермиса, особенно мощным в области вершины когтя, где он постоянно стирается. Роговой слой стенки содержит твердый, слой подошвенной поверхности - мягкий кератин. Под роговым слоем лежит ростковый слой, состоящий из одного ряда базальных клеток и несколько рядов шиповатых клеток. Аналогичную структуру имеет и другое роговое образование - шпора , вырастающая у самцов семейства куриных на плантарном костном отростке плюсно-заплюсневой кости (цевки).
Клюв - роговой чехол надклювья и подклювья. Форма клюва видоспецифична. Наиболее мощный роговой чехол имеют куриные. Он образован роговым слоем эпидермиса, который достигает наибольшей толщины на дорсальной поверхности клюва. По сторонам роговой чехол тоньше. При переходе клюва в слизистую оболочку ротовой полости образуется довольно острый край. У зародышей и птенцов в период вылупления около вершины клюва имеется роговой обизвествленный конусовидный вырост с острой вершиной - яйцевой зуб. Он служит для разрушения скорлупы при вылуплении и вскоре после выхода птенца из яйца отваливается. Роговой чехол клюва, особенно у вершины, постоянно изнашивается и вновь нарастает за счет росткового слоя эпидермиса. Основа кожи имеет незначительную толщину. Она содержит пигментные клетки - меланофоры, поэтому клюв часто пигментирован. Обычно он окрашен в тот же цвет, что и ноги. Клюв - наиболее чувствительное место поверхности тела птицы, в нем имеется большое количество рецепторных окончаний, особенно телец Грандри, Мейснера и Гербста. Корень клюва при переходе в кожу головы покрыт мягкой восковидной кожей - восковицей или церомой, особенно богатой осязательными нервными окончаниями. У гусиных восковица одевает весь клюв и придает ему оранжевый цвет, который в брачный период меняется на красный. По краю надклювья имеются поперечные листочки длиной около 1 мм, заходящие между такими же листочками подклювья. Это рамфотека - фильтр, который задерживает при процеживании планктон, выловленный из воды.
Кожные железы представлены единственной сальной железой - копчиковой. Потовые железы отсутствуют. Копчиковая железа - парная сложная трубчатая сальная железа. Лежит под кожей на свободных хвостовых позвонках в виде двух долей овальной формы, обращенных вершинами назад. Более крупная (длиной 10-15мм) она у водоплавающих и сравнительно небольшая (5-7мм) у сухопутных птиц. У некоторых птиц (страусов, голубей) копчиковая железа отсутствует. Каждая доля железы состоит из большого количества ветвящихся трубочек, которые открываются в общую полость. Стенки трубочек образованы многослойным эпителием и отграничены от прилежащей соединительной ткани базальной мембраной. Базальный слой эпителия является камбиальным. Он состоит из кубических клеток с округлыми, крупными ядрами. В клетках вышележащих слоев увеличивается количество цитоплазмы за счет накопления жировых веществ. Затем клетки превращаются в капли секрета - железа секретирует по голокриновому типу, клетки при этом теряют ядра и органеллы, утолщаются и слущиваются. Секрет вытесняется нарастающими молодыми клетками из просвета трубочек в центральную полость. Центральная полость переходит в сравнительно узкий выводной проток. Стенка выводного протока образована многослойным плоским эпителием. Открывается он на поверхности кожи, на верхушке сосочка, которым оканчивается вершина копчиковой железы. Птица отжимает этот секрет, надавливая клювом на сосочек и перьевую кисточку, растирает по перьям, что предохраняет их от намокания и растрепывания. Соединительная ткань покрывает железу снаружи, отделяет друг от друга железистые трубочки и доли железы, образуя строму органа и междолевую перегородку. Около вершины железы в области перешейка в междолевой перегородке залегают пучки гладкомышечных клеток, которые также способствуют продвижению секрета из центральной полости и по выводному протоку. Секрет копчиковой железы сложного состава. В него входят жиры, белки, нуклеиновые кислоты, много ферментов, и другие биологически активные вещества. Существует мнение, что секрет копчиковой железы отличается от секрета типичных сальных желез тем, что содержит высокоактивные ферменты стероидного метаболизма. Кроме того, считается, что секрет содержит провитамин D - эргостерин, который под действием ультрафиолетовых лучей превращается в витамин D. [7].
2.7.2. Перьевой покров.
Тело птицы покрыто перьями. Однако у большинства птиц перья растут
не по всей поверхности, а на определенных участках, называемых
птирилиями. Они обычно проходят вдоль тела (в направлении полета) у
разных видов птиц по-разному и имеют названия в соответствии с той
частью тела, на которой находятся: головная, шейные, крыловая,
плечевая, боковая, грудная, спинно-поясничная, брюшная, бедренная,
голени, хвостовая, анальная и другие птерилии. Участки тела
свободные от оперения - аптерии. Они так же носят название по
топографическому признаку. Вместе с тем, перья расположены так, что
покрывают аптерии, образуя сплошной перьевой покров. У нелетающих
птиц (страус, пингвин) перья растут по всей поверхности тела.
Назначение перьевого покрова - защита тела от механического
воздействия и одно изприспособлений, обеспечивающих поддержание
температуры тела. В регуляции теплообмена оперение играет большую
роль (пока цыпленок покрыт пухом, разница между температурой кожи и
воздуха составляет 13-15°С, с появлением перьев она достигает
17-19°С). Оперение увеличивает теплоизоляцию, создает вокруг тела
слой неподвижного воздуха, что препятствует теплоотдаче. Изменяя
наклон перьев, птица может регулировать теплоотдачу. Кроме
терморегулирующей функции, перьевой покров создает обтекаемую форму
тела, облегчающую полет, и создает несущие поверхности, делающие
полет возможным. Перья в зависимости от формы и функции делят на:
-
контурные,
-
пуховые,
-
полупуховые,
-
нитевидные,
-
кисточковые,
-
щетинки,
-
порошковый пух (пудровые).
-
Контурные перья - наиболее распространенный тип перьев. Они определяют очертания тела птицы. Среди них различают:
-
кроющие,
-
маховые,
-
рулевые перья.
Кроющие перья расположены на головной и шейной птерилиях. От
боковой, грудной, стернальной и брюшной птерилий отходят кроющие
перья
грудного отдела туловища, хлупа (кончика крестца) и живота. Зрелое
кроющее перо состоит из ствола, стержня и опахала (рис. 8). Нижняя
часть ствола (до опахала) цилиндрической формы называется очином.
Нижняя часть очина заключена в перьевой мешочек и погружена в кожу.
На конце очина есть углубление с отверстием - нижний пупок. В этом
углублении находится зачаток пера следующего поколения. Стенки
очина состоят из довольно прозрачного рогового вещества. У молодого
контурного пера бóльшая часть очина заполнена соединительным
сосочком с богатой кровеносной сетью, которая придает ему красный
или синий цвет. По мере созревания сосочек укорачивается, очин
заполняется воздухом, а ороговевшие клетки эпидермиса, сохраняясь
на прежних местах, превращаются в воронковидные пленки, как бы,
отмечая этапы редукции сосочка, на уровне опахала, постепенно
сужаясь, очин переходит в четырехгранный стержень. В месте перехода
имеется углубление - верхний пупок. От него начинается добавочное
перо.
От стержня в обе стороны под некоторым углом
отходят лучи (бородки) первого порядка, образующие в совокупности
опахало - упругую перьевую пластинку. Самая нижняя небольшая часть
опахала называется шелковистой, средняя - пуховой, верхняя -
контурной. Внешняя половина опахала кроющего пера более узкая и
тугая, внутренняя - более широкая и мягкая. От лучей первого
порядка в обе стороны симметрично под углом отходят многочисленные
лучи (бородки) второго порядка, причем с одной стороны они
располагаются несколько выше, чем с другой. На низлежащих бородках
второго порядка имеется гребень, загнутый в виде карниза, идущего
по верхнему краю луча. Вышележащие лучи второго порядка несут на
себе лучи (бородки) третьего порядка, представляющие собой выросты
ороговевших клеток эпителия в виде ресничек и крючочков. Крючочки
заходят за гребни лучей второго порядка соседнего пера, по которым
они могут скользить, сохраняя упругую связь между собой. Строение
крючочков и их количество видоспецифичны, реснички создают
шероховатость, препятствующую скольжению перьев друг по другу,
увеличивая трение, чем способствуют удержанию перьев в виде
сплошного покрова. Для шелковистой части опахала кроющего пера
характерно рудиментарное состояние лучей второго порядка. В пуховой
части неразвитыми оказываются реснички и крючочки (лучи третьего
порядка). Стержень и лучи первого порядка образованы тремя слоями
клеток. Самый наружный
- кутикула состоит из одного ряда плоского ороговевающего эпителия.
Средний слой - корковый - образован большим количеством рядов
вытянутых и уплощенных ороговевших эпителиальных клеток, плотно
прилегающих друг к другу, что обеспечивает прочность пера.
Внутренний слой - мозговой - состоит из многоядерных крупных
ороговевших клеток, заполненных воздухом, что придает перу большую
легкость. В очине и лучах второго и третьего порядков мозговое
вещество не выражено. Форма клеток и слоев видоспецифична. Кроющие
перья подвижны. Это обеспечивается хорошо равитыми гладкими
мышцами, оплетающими перьевой фолликул и оканчиваются в дерме. При
этом каждое кроющее перо оказывается закономерно связанным с
четырьмя соседними перьями. Такая связь позволяет синхронно
поднимать и опускать кроющие перья тела, мышцы фолликулов.
Прилежащие к ним участки кожи обильно иннервированы как свободными
нервными окончаниями, так и тельцами Гербста.
Рис.8 строение пера.
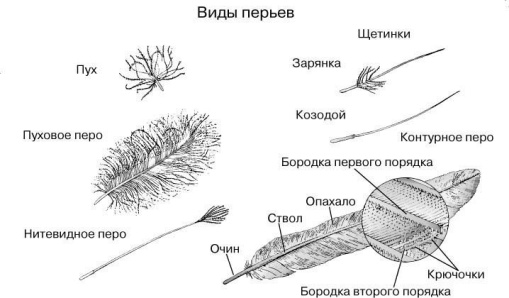
Маховые и рулевые перья устроены аналогично кроющим. В отличие от последних у них длиннее очин, менее развиты шелковистая и пуховая части опахала, и длиннее кроющая часть опахала. Пуховые перья, или пух - мелкие, имеют короткий очин, слаборазвитый стержень и опахало с несвязанными лучами из-за отсутствия крючков и ресничек. Стержень может быть не развит, в таком случае лучи отходят непосредственно от очина, расположен пух на птерилиях между контурными перьями, но может расти и на аптериях, особенно в области хлупа и живота. Основная функция - теплоизоляция, утепление тела птицы. Наиболее развит пух у водоплавающих. Полупухвые перья - имеют такое же строение, как и пух, но стержень у них всегда развит. распространены перья этого типа в области хлупа и живота. Нитевидные перья - имеют длинный, очень тонкий стержень, на вершине которого находится маленькое опахало, состоящее всего из нескольких соединенных лучей. Расположены они в количестве 1-8, всегда у контурного пера, прикрыты им и развиваются, по-видимому, из одного с ним сосочка. У индюка растут на груди в виде пучка. В сумках нитевидных перьев обнаружены многочисленные нервные окончания. Считается, что они обладают рецепторной функцией, с их помощью птица ощущает непорядок в
Контурном пере и устраняет его. Возможно, от них
происходит сигнализация, необходимая для управления движением
оперения. При этом очевидно, рефлекторная дуга замыкается на мышцах
контурного пера. Кисточковые перья имеют тонкий ствол и слабо
сцепленные лучи, расходящиеся наподобие волосков кисти,
располагаются вокруг протока
копчиковой железы. Щетинки - короткие перышки, состоящие из
небольшого ствола без опахала. Они бывают у основания клюва -
вибрисы, около ноздрей и глаз. Порошковый пух (крошащийся,
пудровый) хорошо развит у птиц, у которых копчиковая железа
отсутствует или слабо развита (голуби, цапли и др.). [6].
2.8. Аппарат Пищеварения.
Аппарат пищеварения принято разделять на четыре отдела, которые отражены в нижеследующей рис. 9.
Рис. 9
Деление пищеварительного тракта птицы на отделы.
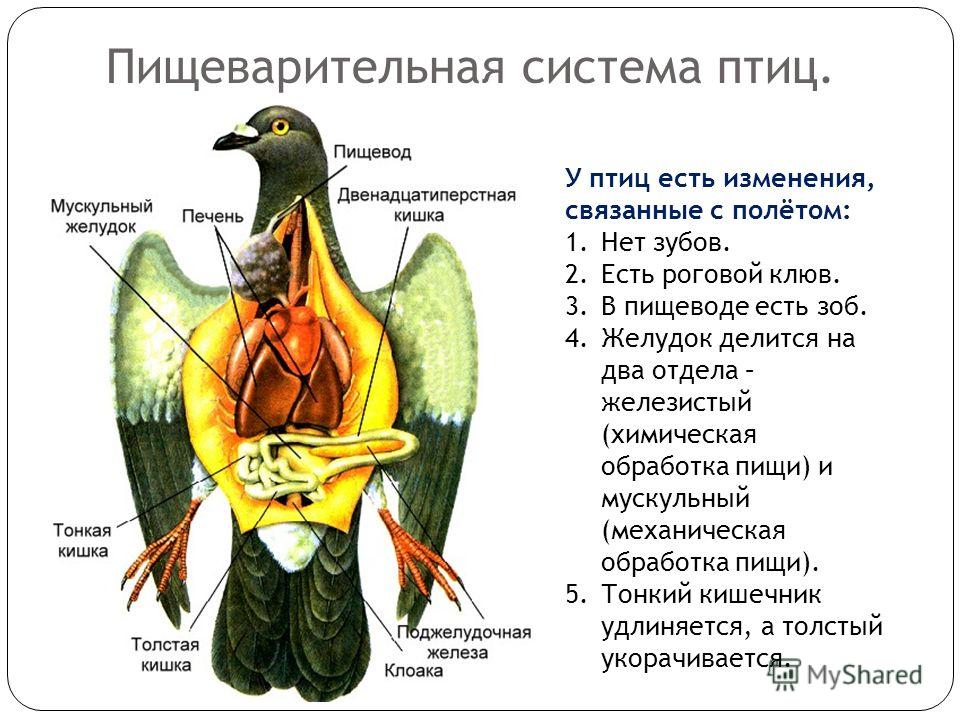
Строение пищеварительной системы птиц во многом напоминает пищеварительный аппарат млекопитающих. Она включает ротоглотку, пищеводно-желудочный отдел, тонкий и толстый кишечник. Характер переработки твердого корма, поступающего в организм птиц и требующего измельчения при отсутствии зубов, обуславливает особенности функциональной морфологии пищеварительной системы. Значительные отличия имеют ротовая полость и желудок. Другие органы - толстая и тонкая кишка, печень, поджелудочная железа сходны по строению с органами млекопитающих.
Внутреннее строение птиц рис. 10
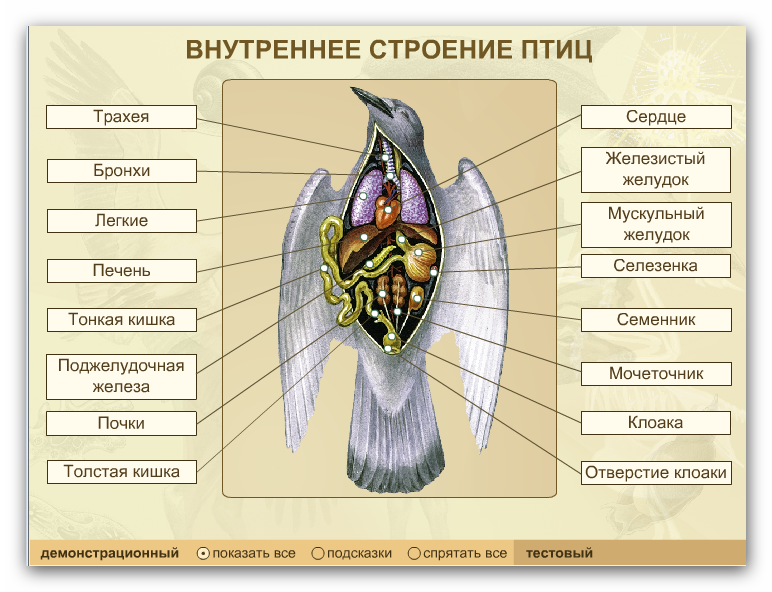
В состав ротоглотки или головной кишки входит
ротовая
полость и глотка, которые у птиц не отделены друг от друга из-за
отсутствия
небной занавески. Кроме того, у птиц отсутствуют губы, щеки, и
десны;
челюсти преобразовались в клюв. В результате отсутствия
перечисленных
органов у птиц нет преддверия ротовой полости. Собственно ротовая
полость
- пространство, ограниченное спереди и с боков клювом, сверху -
твердым
небом, снизу - дном. Задняя граница между ротовой полостью и
глоткой
проходит по заднему ряду небных сосочков и сосочков языка.
Костной
основой ротовой полости являются кости черепа.
Ротовая полость птиц покрыта плоским многослойным эпителием. Толщина эпителиального слоя и характер его ороговевания в разных участках ротовой полости характеризуется своими особенностями. Процесс ороговевания интенсивно протекает на границе с кожным покровом и завершается образованием кожного пласта. Многослойный плоский орговевающий (сквамозный) эпителий расположен на основной пластинке, построенной из рыхлой соединительной ткани. Крышей ротовой полости является твердое небо. У кур оно имеет узкую небную щель, поперек небной щели размещены пять видов сосочков. На твердом небе и по бокам от него расположены отверстия слюнных желез, кроме того, небольшие слюнные железы расположены по бокам от средней и задней части языка, и на дорсальной поверхности основания языка. У птиц дополнительно имеются железы угла рта и передние, и задние подчелюстные железы. В основе строения этих желез лежит железистая долька, состоящая из слизистых клеток, радиально расположенных вокруг собирательного или центрального пространства (полости). Последняя переходит в выводной проток, стенка которого построена покровным эпителием. Серозные секреторные концевые отделы в составе слюнных желез не обнаружены. Строение слюнных желез птиц изучено не достаточно. Снизу ротовая полость ограничена языком. Различают в языке верхушку, тело, соединенное уздечкой с дном ротовой полости, спинку, боковые части и корень. Основу языка составляет плотная соединительная ткань, содержащая эластические волокна. Снаружи язык покрыт слизистой оболочкой с сильно ороговевающим многослойным плоским эпителием. На аборальном конце спинки слизистая образует поперечный ряд высоких нитевидных сосочков. В собственной пластинке слизистой оболочки залегают пакеты сложных трубчатых слизистых желез, выводные протоки, которых открываются на спинке, боковых частях и корне языка. С выводными протоками тесно связаны чувствительные вкусовые рецепторы - вкусовые почки, залегающие в эпителиальном пласте. По строению напоминают вкусовые луковицы млекопитающих. Количество их незначительно. В языке имеются тактильные, болевые нервные окончания. Поперечно-полосатая мышечная ткань формирует три небольшие язычные мышцы. Язык птиц прочно соединен с подъязычной костью. Составная часть подъязычной кости заходит внутрь корня языка. Все остальные мышцы, приводящие в движение язык, относятся к подъязычному аппарату.
Ротовая полость переходит в глотку - участок
кишечной трубки, где перекрещиваются дыхательные и пищеварительные
пути. На дне глотки имеется большое овальное отверстие - вход в
гортань. Позади его, на границе между дном глотки и пищеводом,
расположен поперечный ряд конусовидных гортанных сосочков. Стенка
глотки состоит из слизистой, мышечной оболочек и адвентиции.
Слизистая покрыта
многослойным плоским эпителием. Собственная пластинка слизистой
оболочки образована рыхлой соединительной тканью, в которой
залегают железы, имеются лимфатические и сосудистые сплетения.
Лимфатические сплетения формируют глоточные миндалины.
В состав переднего (пищеводно-желудочного)
отдела входят пищевод с
зобом и двухкамерный желудок, состоящий из железистой и мышечной
частей (рис. 10). Пищевод (esophagus) - трубкообрзный орган,
начинающийся за глоткой. В начальном участке пищевод лежит над
трахеей, ближе к входу в полость тела он переходит в правую сторону
и располагается справа от трахеи. Каудальная часть пищевода вновь
оказывается над трахеей, проходит между бронхами, легкими, над
сердцем и без резких границ, слегка сужаясь, переходит в железистый
желудок. Длина пищевода у кур 25-30 см. У куриных около входа в
полость тела пищевод мешкообразно расширяется образуя зоб. Стенка
пищевода состоит из слизистой, мышечной и адвентициальной оболочек.
В слизистой оболочке хорошо выражены четыре слоя: эпителиальный,
основная и мышечная пластинки, подслизистая основа. Эпителиальный
слой представлен плоским многослойным ороговевающим эпителием. В
процессе орговения поверхностно лежащие клетки превращаются в
роговые чешуйки. Собственная пластинка состоит из рыхлой
соединительной ткани, которая бедна эластическими волокнами и
лимфоидной тканью. Ее сосочки вдаются в эпителиальный слой и имеют
значительную высоту. В самой пластинке расположены слизистые
железы. Все концевые отделы железы открываются в собирательную
полость, являющуюся началом неразветвленного протока. В железистых
клетках концевого отдела ядра прижаты слизистым секретом к
базальной мембране. В зоне перехода основной пластинки пищевода в
желудок лежат многочисленные лимфоидные узелки - эзофагиальная
миндалина.
Мышечная пластинка слизистой оболочки сильно
развита. Она построена из продольно ориентированных гладкомышечных
клеток. Этот слой участвует в образовании складок слизистой
оболочки. Подслизистая основа состоит из рыхлой соединительной
ткани, создавая тем самым подвижность слизистой оболочки при
образовании ее непостоянных складок. Мышечная оболочка представлена
двумя слоями гладкомышечных клеток:
внутреннего - циркулярного, наружного - продольного. Более развит
циркулярный слой.
Адвентиция, как и у млекопитающих, построена из рыхлой соединительной ткани. После вхождения пищевода в грудную полость адвентиция заменяется серозной оболочкой. Производным стенки пищевода является зоб, поэтому его стенка так же имеет три оболочки, построены из тех же слоев, а слои из тех же - тканей.
Вентральная стенка зоба представлена более толстым эпителиальным слоем, в котором очень четко проходит граница между производящим и роговым слоями. Слизистые железы находятся только в дорсальной стенке зоба. Мышечная пластинка и мышечная оболочка особенно сильно развиты в вентральной части.
Пищеварение в зобе идет за счет ферментов корма, микроорганизмов и амилолитических ферментов слюны. В 1г содержимого зоба насчитывается до 0,1 млрд. микроорганизмов, в основном аэробов, обнаружены грибки и дрожжевые клетки.
У самцов и самок голубей в период кормления птенцов в зобе появляется жирная белая масса - «зобное молочко», которая образуется в результате разрастания и перераспределения слизистых оболочек зоба. Отрыгивая «зобное молочко», голуби кормят им птенцов в первые 10-16 днейжизни. В его состав входят 16% белка, 11% жира, 1,3 минеральных веществ, витамины А и группы В. [3].
Желудок (ventriculus s. gaster) - состоит из двух отделов - железистого и мышечного (рис. 8). Первый выделяет пищеварительный сок, второй предназначен для перетирания пищи. В железистом желудке пищевой ком обогащается ферментами и, не задерживаясь, попадает в мышечный желудок, где происходит химическая и механическая переработка. Его стенка сформирована из слизистой, мышечной и серозной оболочек. Эпителиальным слоем слизистой оболочки является однослойный цилиндрический железистый эпителий, поэтому покровный эпителий желудка - это обширное железистое поле, продуцирующее слизь. Основная пластинка представлена рыхлой соединительной тканью, богатой клеточными элементами.
Выводные протоки желез открываются на
поверхности возвышений
слизистой оболочки. Эти возвышения видны невооруженным взглядом и
называются железистыми мешочками. Других желез в слизистой оболочке нет. Мощный слой мышечной пластинки слизистой оболочки железистого желудка являетсяпродолжением мышечной пластинки слизистой пищевода. Гладкомышечные клетки оплетают железы снизу, с боков и сверху. Подслизистая основа состоит из рыхлой соединительной ткани и развита незначительно. Мышечная оболочка представлена двумя слоями гладкомышечных клеток, из которых внутренний является циркулярным, наружный - продольным. Серозная оболочка построена из рыхлой соединительной ткани и мезотелия.
Кишечник подразделяется на толстый и тонкий
отделы. Тонкий кишечник (intestinum tenue). Состоит из
двенадцатиперстной, тощей и подвздошной кишок.Следует обратить
внимание на хорошо развитые застенные железы: печень, состоящую из
двух долей; и поджелудочную железу, залегающую в петле
двенадцатиперстной кишки. Процессы всасывания у птиц интенсивнее
идут в тощей кишке. По строению стенка кишечника сходна со
строением млекопитающих. Эпителиальный слой слизистой оболочки -
однослойный цилиндрический каемчатый эпителий. Он состоит из
каемчатых, бокаловидных и энтерохромаффинных клеток. Основная
пластинка построена из рыхлой соединительной ткани, образует
выпячивания, покрытые каемчатым эпителием - это ворсинки (на всем
протяжении кишечника). Лимфатическая система ворсинок слабо
развита, вследствие чего жир всасывается непосредственно в кровь. У
основания ворсинок открываются крипты - трубкообразные вдавления
так же покрыты эпителием. Они увеличивают поверхность всасывания, у
их основания находятся железистые и стволовые клетки. Поэтому
крипты считаются зоной
митотически делящихся энтероцитов, которые восполняют эпителиальный
слой ворсинок. Соединительная ткань богата лимфоидными элементами,
которые расположены диффузно в виде лимфоидных узелков.
Подслизистая основа тонкого кишечника образована рыхлой и ретикулярной тканями, дуоденальные железы в подслизистой основе двенадцатиперстной кишки у птиц отсутствуют. На границе с мышечной оболочкой находится подслизистое (Мейснерово) нервное сплетение.
Печень (hepar) крупная, состоит из двух долей. У цесарки, страуса и голубя нет желчного пузыря. Функциональная морфология птиц и млекопитающих сходна: дольчатое строение обусловлено кровоснабжением органа; центральное расположение в дольке центральной вены; на периферии долек находятся триады, состоящие из междолькового желчного протока, междольковой вены и артерии; гепатоциты формируют радиально лежащие балки. Между ними находятся венозные синусоиды.
Поджелудочная железа (pancreas) - дольчатый
орган. Количество железистых долек у сельскохозяйственных птиц
разных видов варьирует. Дольки разграничены междольковой
соединительной тканью, они построены из экзокринных и эндокринных
отделов. Главные выводные протоки распадаются на междольковые, а
междольковые в свою очередь - на вставочные. Главные и междольковые
выстланы однослойным цилиндрическим эпителием, вставочные -
однослойным плоским. Эндокринная часть железы - комплекс
эндокринных островков Лангенгарса, выделяющих гормон инсулин. У
птиц они состоят только из одного типа клеток - либо клеток А
(темных), либо из клеток В (светлых), в связи с чем различают
темные и светлые островки. Соотношение этих типов островков может
меняться под влиянием пола и возраста. Толстый кишечник (intestinum
crassum). Этот отдел кишечника укорочен, в нем выделяют две слепые
кишки и одну прямую, которая впадает в клоаку (рис. 10).
Пищеварение в слепых кишках птицы происходит с помощью ферментов,
поступающих с химусом из тонкого отдела кишечника, и микрофлоры.
Здесь происходит сбраживание клетчатки и незначительное ее
переваривание (6-9%). В слепых кишках происходит всасывание воды,
минеральных веществ, продуктов брожения, синтез витаминов группы В,
в
том числе витамина В12.
Стенка толстого кишечника состоит из слизистой с подслизистой основой, мышечной и серозной оболочек. Слизистая оболочка имеет такое же строение, как и в тонком отделе:характерны ворсинки и общекишечные железы. В толстом отделе кишечника увеличивается содержание бокаловидных клеток, наблюдается значительное развитие лимфоидных образований (особенно в слепых кишках). Остальные оболочки сходны по строению с оболочками тонкого кишечника. Каудально прямая кишка расширяется и переходит в клоаку. В нее открываются половые и мочевые пути, поэтому в ней различают три отдела: копродеум, уродеум, проктодеум. Первый отдел является самым обширным. По строению клоака имеет сходство с задней кишкой. В проктодеум открывается клоакальная (фабрициева) сумка, которая является эпителиально лимфатическим органом иммунологической защиты организма. [6].
2.9. Аппарат дыхания.
Органы дыхания у птиц имеют ряд особенностей:
- малая величина и несложность строения носовой полости
- наличие в бифуркации трахеи приспособления для издания звука-
певчей
гортани
- незначительная величина и положение легких, бронхи которых
сообщаются
с полостями воздухоносных мешков.
В носовой полости имеются три хрящевые раковины, отсутствует лабиринт решетчатой кости. Через небную щель из носовой полости воздух попадает в ротоглотку, которая так же, как у млекопитающих, сообщается слуховыми трубами со средним ухом. Верхняя гортань не имеет надгортанника и щитовидного хряща. Голосовой (певчая гортань) аппарат расположен на бифуркации трахеи. Трахея (thrachea) - полая зияющая трубка длиной 16-27 см., лежащая в области шеи и передней части полости тела. Стенка трахеи содержит три оболочки: слизистую, фиброзно-хрящевую и адвентицию. В состав слизистой входит однослойный многорядный мерцательный эпителий, включающий бокаловидные клетки. Подэпителиальный соединительнотканный слой слизистой оболочки трахеи состоит из рыхлой неоформленной соединительной ткани и имеет слизистые трахеальные трубчатые железы. В полости тела перед впадением в легкие трахея делится на два главных бронха - бифуркация.
В каждое легкое птицы поступает один главный
бронх. В паренхиме органа он распадается на четыре вторичных
бронха. Последние, вновь разветвляясь, образуют более тонкие
третичные бронхи или парабронхи. Легкое (pulmones) птиц содержит
парабронхи, окружающую их паренхиму органа, разделенную на нерезко
отграниченные дольки. Диаметр парабронхов у курицы составляет
100-150мкм. Их стенка состоит из циркулярно ориентированных гладких
мышечных клеток и эластических волокон. Внутренняя поверхность
парабронхов выстлана однослойным кубическим или плоским эпителием.
Дольки легкого представлены воздухоносными капиллярами,
принимающими воздух из парабронхов, кровеносными капиллярами и
соединительной тканью. Воздухоносные капилляры по строению и
функциональному значению соответствуют легочным альвеолам
млекопитающих и отличаются тем, что у птиц в легких, а
соответственно и в воздухоносных капиллярах газообмен происходит
при вдохе и выдохе. Воздухоносные капилляры образуют сеть в
пределах одной дольки, но могут анастомозировать и с воздухоносными
капиллярами соседних долек. Воздухоносные капилляры птиц - трубки
неправильной формы с диаметром от 2 до 6 мкм. Их стенка состоит из
базальной мембраны и одного слоя очень плоских эпителиальных
клеток. Наиболее толстая часть последних около ядра достигает
0,3-0,5 мкм, тогда как по периферии - 20-40 нм и там нет органелл и
микроворсинок. Базальная мембрана беструктурна. Такое же строение
имеют капилляры, соединяющие межкапиллярные воздухоносные протоки,
но их просвет еще и уже, часто не спадается совсем. Воздухоносные и
кровеносные капилляры разделяет лишь субмикроскопическое
пространство в 60-100 нм, заполненное плотным гомогенным веществом.
Барьер между воздухом и кровью не достигает 1 мкм. При вдохе и
выдохе птиц изменяется преимущественно объем воздухоносных мешков,
а не легких, вентиляция которых, а соответственно
всей системы органов дыхания - результат сокращения грудной и
брюшной стенок.[2].
Легкие птиц небольшие по объему, не делятся на доли, глубоко
проникают в межреберные промежутки, располагаются от первого ребра
до
почки. С легкими птиц связано 5 пар воздухоносных мешков.
Функции воздухоносных мешков разнообразны:
- они являются дополнительными резервуарами воздуха;
- повышают газообмен;
- участвуют в терморегуляции и водном обмене;
- облегчают массу тела;
- выполняют роль резонаторов, амортизаторов и теплоизоляторов.
Воздухоносные мешки представляют собой внелегочное выпячивание воздухоносных путей. Стенка их состоит из соединительной ткани, выстланной с внутренней стороны однослойным кубическим или цилиндрическим эпителием, а снаружи или серозной оболочкой, или адвентицией. [3].
3. Аппарат мочевыделения.
Мочевыделительная система состоит только из
почек и мочеточников,
открывающихся в уродеум клоаки. Лоханка, мочевой пузырь,
мочеиспускательный
канал у птиц отсутствуют. Функция мочевыделительной системы состоит
в удалении избытков воды и солей из организма и поддержания тем
самым постоянства осмотического давления в тканях тела.
Почки у птиц относительно большие, по сравнению
млекопитающими, из-за более интенсивного обмена веществ и
отсутствия потовых желез. Они продолговатой формы, расположены в
вентральных углублениях пояснично-крестцового отдела позвоночника и
подвздошной кости. Окружены воздухоносными мешками, функционально
заменяющими
жировую подушку, отсутствующую у птиц. Капсула почки делится на три
доли: переднюю, среднюю и заднюю.
Почка - компактный орган. Имеется строма (соединительнотканная капсула, междольковые прослойки, делящие почки на дольки) и нефрон (основная структурная и функциональная единица почки).
Нефроны обладают сособностью к значительной реабсорбции воды и растворенных в ней веществ. Почка функционально и морфологически тесно связана с кровью. В отличие от млекопитающих, в почку входят не только артерии, но и вены, образуя воротную систему почки. Артериальная кровь поступает по почечным артериям, которые в паренхиме разветвляются на междольковые и внутридольковые артерии. От последних отходят приносящие артериолы несущие кровь к капиллярам сосудистых клубочков, которые образуют чудесную сеть с капиллярами выносящих артериол. На периферии дольки выносящая артериола соединяется с венозными капиллярами воротной системы почки, оплетающими канальцы нефрона. [2].
4. Аппарат размножения.
Система органов размножения обеспечивает продолжения вида. Эта система состоит из половых желез (семенников или яичников), в которых образуются половые клетки, и проводящих половых путей, по которым зрелые половые клетки передвигаются к органам совокупления. В результате совокупления самца и самки появляется возможность оплодотворения - слияния спермия с яйцеклеткой, образования зиготы и развитие нового организма. Половые железы кроме того, являются железами внутренней секреции, вырабатывающими половые гормоны. Они определяют половое поведение птиц и их внешний облик, так как способствуют развитию вторичных половых признаков. Функция половой системы находится под контролем центральной нервной системы и регулируется гипоталамо-гипофизарной системой.
Мужские гонады - это пара семенников, в которых формируются мужские половые клетки (гаметы) - сперматозоиды. Форма семенников овальная или эллиптическая, причем левый обычно крупнее. Они лежат в полости тела вблизи переднего конца каждой почки. Перед наступлением сезона размножения стимулирующее действие гормонов гипофиза заставляет семенники увеличиваться в сотни раз. По тонкой извитой трубке, семявыносящему протоку, сперматозоиды попадают из каждого семенника в семенной пузырек. Там они накапливаются до наступающего в момент копуляции семяизвержения, при котором выходят в клоаку и через ее отверстие наружу.
Женские гонады, яичники, образуют женские гаметы - яйцеклетки. У большинства птиц яичник только один, левый. По сравнению с микроскопическим сперматозоидом яйцеклетка огромна. Основную ее часть по массе составляет желток - питательный материал для развивающегося после оплодотворения зародыша. Из яичника яйцеклетка попадает в трубку, называемую яйцеводом. Мышцы яйцевода проталкивают ее мимо различных железистых областей в его стенках. Они окружают желток белком, подскорлуповыми оболочками, твердой, содержащей кальций скорлупой и, наконец, добавляют окрашивающие скорлупу пигменты. Превращение яйцеклетки в готовое к откладке яйцо занимает ок. 24 ч. [3].
Оплодотворение у птиц внутреннее. Сперматозоиды попадают во время копуляции в клоаку самки и плывут вверх по яйцеводу. Оплодотворение, т.е. слияние мужской и женской гамет, происходит в верхнем его конце прежде, чем яйцеклетка покроется белком, мягкими оболочками и скорлупой.
5. Сердечно-сосудистая система.
Сердце (cor) у птиц, как и у млекопитающих,
четырехкамерное. В правом желудочке нет сосочковых мышц, правое
атривентрикулярное отверстие щелевидное, оно прикрывается сильно
развитой мышечной пластинкой. У птиц развивается правая дуга аорты.
Брюшная аорта, давая наружную и внутреннюю подвздошную артерии,
делится на две седалищных
(правую и левую) и среднюю крестцовую артерии. У птиц имеются две
краниальные полые вены, а также две воротные системы: печени и
почек. Лимфатическая система у птиц имеет ряд особенностей. Она
представлена малочисленными лимфатическими узлами. Лимфа собирается
в два лимфатических грудных протока: левый и правый,
анастомозирующие друг с другом. Селезенка у птиц овальная либо
круглая, расположена на правой стороне желудка. [6].
6. Нервная система.
Нервная система - интегрирующая и регулирующая
система. По топографическим признакам ее делят на центральную и
периферическую. К центральной относят головной и спинной мозг, к
периферической - ганглии, нервы, нервные сплетения и
нервныеокончания. В связи с особенностями строения и
функционирования нервную систему делят на соматическую и
вегетативную. Соматическая нервная система иннервирует, в основном,
скелет, мышцы, кожу, а также осуществляет связи с внешней средой.
Вегетативная - внутренние органы, эндокринную и
сердечно-сосудистую
системы, регулирует обмен веществ в организме. У птиц аналогичное с
млекопитающими строение спинного мозга, спинномозговых нервов и 12
пар черепно-мозговых нервов. Спинной мозг имеет более выраженное
пояснично - крестцовое утолщение. Особенностью утолщения птиц
является наличие в нем нейроглиального, богатого гликогеном ядра
неизвестной функции - гликогенового тела, которое значительно
увеличивает размеры утолщения.
Латеральные рога в сером веществе спинного мозга у птиц не развиты. Центры симпатической нервной системы расположены вокруг центральной части спинномозгового канала. Из 12 пар черепно-мозговых нервов особенно отличается VII пара - лицевой нерв. Он слабо выражен в связи с отсутствием мимической мускулатуры. Возвратный нерв, идущий в начале X пары, иннервирует, главным образом, пищевод и зоб. Сам блуждающий нерв у птиц развит сильно, XII пара - подъязычный нерв, кроме мышц языка и подъязычной кости, иннервирует певчую гортань. Головной мозг у птиц так же подразделен на пять отделов (рис. 11) - латеральная поверхность. [7].
Головной мозг с латеральной стороны. Рис.11
Каждый отдел головного мозга имеет свои
особенности: 1. конечный мозг - плащ полушарий не имеет извилин,
нет мозолистого тела, на дне боковых желудочков нет аммоновых
рогов, нет полупрозрачной перегородки; 2. промежуточный мозг -
зрительные бугры гораздо меньше, нет сосцевидного тела; 3. средний
мозг - сильно развито двухолмие. Сильвиев водопровод образует
широкую полость;
4. Задний мозг - мозжечок почти не имеет полушарий, червячок
сильно
выражен; 5. Продолговатый мозг выпукл и образует вентральную
кривизну. 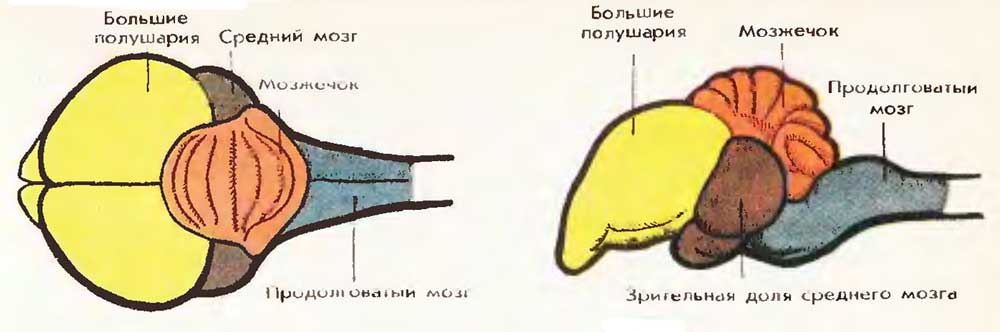
Выводы.
1. Разработанный в настоящей работе системный
морфо-функциональный/морфо - экологический подход показал себя
эффективным орудием в изучении и реконструкции эволюции и
адаптивных преобразований аппарата двуногой локомоции птиц,
способным формировать новые представления о специфике локомоторных
адаптаций
исследуемых групп птиц. Предлагаемый системный подход предполагаем
анализ некоторых наиболее значимых для реконструкции адаптивной
эволюции задних конечностей птиц структур, каковыми являются
компоненты расширенной мускульной формулы и типы взаимодействия
конечных сухожилий длинных глубоких сгибателей пальцев.
2. Типы взаимодействий конечных сухожилий длинных глубоких сгибателей пальцев стопы птиц служат ценным источником информации об адаптивной эволюции аппарата двуногой локомоции птиц и позволяют высказывать суждения об их филетической эволюции.
3. Системный подход к анализу особенностей строения задних конечностей водных птиц позволил показать, что они, подобно другим своим соседям по отряду, имели укороченный палец, который пришлось восстанавливать при переходе в новую адаптивную зону (передвижение по плавающей растительности).
4. Удлинение когтя первого пальца шпорцевых
кукушек связано с адаптацией к передвижению по «ковру» из спутанной
растительности. Конфликт, возникший между удлиненным за счет когтя
первым пальцем и необходимостью охвата насеста стопой, был разрешен
за счет сходного с водными птицами, изменения конфигурации конечных
сухо-
жилий длинных глубоких сгибателей пальцев стопы.
5. В строении задних конечностей птицы сохраняются следы более древних состояний, которые поддаются расшифровке в форме реконструированных ключевых адаптаций и процесса смены в конкретных филетических линиях (Кукушкообразные, Ракшеобразные, Трогонообразные, Дятлообразные, Воробьинообразные).
6. Системный подход к изучению особенностей строения задних конечностей из отряда трогонов показал, что в своей эволюции они прошли через козодоеподобную стадию с ключевой адаптацией к питанию плодами, срываемыми с подлета.
7. Естественная группа Кукушкообразных включает кукушек, отряд тураковых и гоациновых. Последний строением задних конечностей напоминает гипотетическую предковую форму отряда, хотя уже несет некоторые черты специализации к поперечному охвату насеста. Бегающие кукушки Нового и Старого Света несут в строении задних конечностей черты древесной специализации предка.
8. Системный подход к анализу задних конечностей, несмотря на разнообразие локомоторных адаптаций рецентных групп Кукушкообразных, указывает на их первоначальную ключевую локомоторную адаптацию - бег в ветвях крон с поочередной опорой на каждую конечность.
9. Особая конфигурация конечных сухожилий
длинных глубоких сгибателей пальцев стопы птиц у Дятлообразных
указывает на их совместное с Ракше- и Трогонообразными
происхождение из группы анцестральных корацииморфных видов и не
связана с исходной адаптацией к лазанию по стволам деревьев.
Базовый облик сухожильной системы стопы Дятлообразных связан с
приобретением зигодактилии и не имеет от-
ношения к предлагаемой рядом авторов адаптации предка всего отряда
к карабканию по стволам деревьев.
10. Морфология задних конечностей показывает, что ключевая локомоторная адаптация Воробьинообразных заключается в передвижение в кронах прыжками с исключительной опорой на обе конечности, конструкция которых в связи с этим существенно упрощена в сочетании с высокой энергетикой обмена.
11. Задние конечности археоптерикса, хотя и могут служить иллюстрацией некоего промежуточного этапа на пути к аппарату двуногой локомоции современных птиц, все же напоминают таковые манираптор и адаптированы к лазанию по стволам деревьев (без опоры на хвост) и бегу по грунту и ветвям. Для поперечного охвата насеста они практически не использовались.
12. Аппарат двуногой локомоции птиц в своем формировании прошел через ряд адаптивных стадий. Приобретя базовые черты в ходе бипедальной наземной локомоции, она модифицировался при карабкании предковых форм (близких к археоптериксу) по стволам деревьев и получил нынешнюю конфигурацию в ходе адаптации передних конечностей к активному полету при одновременной редукции хвоста.
Список литературы.
1. Зиновьев А.В., 2004. К вопросу о происхождении гетеродактилии у
трогонов (Aves: Trogoniformes) // 4 науч. чтения памяти проф.
В.В.Станчинского. Смоленск: СмГУ. С. 374-378.
2. Зиновьев А.В., 2007. Случаи необычной конфигурации конечных сухожилий длинных сгибателей пальцев в двух группах птиц: морфо-функциональный анализ и эволюци- онное приложение // Бюл. Моск. об-ва испыт. природы. Т. 112. Вып. 5. С. 3-8.
3. Прам Р., Баш А. Динозавры или птицы: кто оперился первым? - truemoral.ru/peria.html // В мире науки. - 2003. - № 7. (Статья об эволюционном происхождении перьев.)
4. Френсис П. Птицы. Полная иллюстрированная энциклопедия // Москва: АСТ - Дорлинг Киндерсли, 2008. - 512 с. - (Дорлинг Киндерсли).
5. Бейчек В., Штястны К. Птицы. Иллюстрированная энциклопедия // Москва: Лабиринт-пресс, 2004. - С. 69. - 289 с.
6. Михаил Калякин, Н. Зеленков, В. Гаврилов, 2013. Проблемы эволюции птиц. Систематика, морфология, экология и поведение. Материалы международной конференции памяти Е. Н. Курочкина // на базе Звенигородской биологической станции имени С.Н.Скадовского биологического факультета МГУ, 224с.
7. Джон Вудкок</<font size="4">. Птицы. Полная иллюстрированная энциклопедия // Москва: АСТ- 2015.- 512 c.
54