- Учителю
- Ғылыми жұмыс Тақырыбы: «Petunia Juss. (Петуния) туысы түрлерін in vitro әдісімен көбейту»
Ғылыми жұмыс Тақырыбы: «Petunia Juss. (Петуния) туысы түрлерін in vitro әдісімен көбейту»
Н. Құлжанова атындағы Торғай гуманитарлық колледжі, Арқалық қ.,
Қазақстан Республикасы
Ғылыми жұмыс
Тақырыбы: «Petunia Juss. (Петуния) туысы түрлерін in vitro әдісімен көбейту»
Жұмысты орындаған:
Оқытушы Сапарова А.А.
Арқалық, 2016
МАЗМҰНЫ
БЕЛГІЛЕР МЕН ҚЫСҚАРТУЛАР.......................................................... 3КІРІСПЕ……………………………………………………….................
4
І. ӘДЕБИЕТКЕ ШОЛУ
1.1.Клеткаларды іn vіtro жағдайында өсірудің жалпы сипаттамасы.........
7
ІІ. ЗУРТТЕУ ӘДІСТЕРІ
2.1.
In vitro өсіретін өсімдіктерді залалсыздандыру жұмыстарын жүргізу әдістері....................................................................................
16
2.2
Клеткалардың қоректенуі және қоректік орталардың жалпы сипаттамасы..........................................................................................
18
2.3.
Өсімдік материалын дайындап, эксплаптты бөліп алу әдісі...........
29
2.4.
Эксплантты агары бар ортаға отырғызу және өсіру әдісі......................
32
2.5.
Асептикалық өскіндерден каллус ұлпаларын алу әдісі...........................
34
2.6.
Өсімдіктерді микроклонды көбейту және отырғызу әдісі......................
35
ІІІ. КҮТІЛЕТІН НӘТИЖЕЛЕР
3.1.
Лабораториялық жағдайда Petunia Juss. гүлін in vitro әдісімен көбейту......................................
39
ҚОРЫТЫНДЫ………………………………………......................
ПАЙДАЛАНЫЛҒАН ӘДЕБИЕТТЕР……………...........….......
40
41
БЕЛГІЛЕУЛЕР МЕН ҚЫСҚАРТУЛАР
НТИ - нақты температуралық инактивация
РНҚ - рибонуклеин қышқылы
ДНҚ - дезоксирибоза нуклеин қышқылы
ИСҚ - индолсірке қышқылы
рРНҚ- рибосомалық РНҚ
ЭДТА - этилендиаминтетрасірке қышқылы
тРНҚ- тасымалдаушы РНҚ
мРНҚ- матрицалық РНҚ
ГҚ- гиббереллин қышқылы
АБК-Абсциз қышқылы
Кіріспе
Тақырыптың өзектілігі: Қазіргі кезде өсімдік жасушалары мен ұлпаларын өсіруге айтарлықтай көп көңіл бөлінуде. In vitro жағдайында өсімдіктерді өсіру іргелі зерттеулер болып табылатын өсімдіктер генетикасы, физиологиясы, цитологиясы және молекулалық биология сияқты ғылым салаларын дамыта отырып сәндік өсімдіктердің өнімділігін жоғарылатып сапасын көтеруде маңызды рөл атқарады.
Гүл өсіру - бақтарда, саябақтарда, саяжайларда, ғимараттарды әшекейлеуде, қаланы көгалдандыруда маңызды іс.
Қаланы көгалдандыру жұмыстарын жүргізуде бір, екі не көп жылдық және кілем тәрізді өң беретін өсімдіктер өсіріледі. Ол үшін өсімдік түрлерінің биологиялық ерекшеліктері, гүлінің әсемдігі ғана емес, гүлдің түсі, пішіні, жапырақтарының үлкен-кішілігі, сабағының ұзындығы, гүлдеу мерзімі, ұзақтығы, өсімдіктердің бір-бірімен үйлесімділігі ескеріледі.
Ғылыми деректер бойынша, белгілі бір аймақтың 15%-ы көгалдандырылса, шаң-тозаңның мөлшері 20%-ға кемиді, 35%-на өсімдік өсірілсе, шаң 50%-ға азаяды, ал 65%-ға өсірілсе, шаң 95%-ға кемиді. Жол жағалауларына отырғызылған ағаш пен бұталар, гүлдер мен көгал ауадағы көмір қышқыл газды сіңіріп, ауаны отгегімен байытады, шуды азайтып, мекен жайды желден, күннен және шаңнан қорғайды. 1 га жасыл алқап 1 сағатта ауадан 8 кг көмірқышқыл газын бойына сіңіреді. Көгалдандыру жұмыстарын жүргізгенде міндетті түрде көгалдандырылатын алқаптың ауа райы, топырақ құрамы, жер бедері сияқты ерекшеліктері ескеріледі, суаруға қажетті су көздері қарастырылады. Осы факторлар бойынша Арқалық қаласының климаттық жағдайына төзімді in vitro әдісімен Petunia туысы түрлерінің санын тез көбейтіп, ғимараттардың ауласын, саябақтарды безендіруге үлес қосуға болады.
Өсімдіктердің мүшелері мен жекелеген бөліктерін өсіруді алғаш рет ХІХ-ХХ ғасырда неміс ғалымдары бастаған. Карл Рехингер 1883 жылы алғаш рет бүршіктерді, тамыр және сабақ үзіндісін өсірді. Ол каллустың түзілуін байқады, бірақ қоректік ортаны дұрыс таңдамауынан ұзақ уақыт дақылды өсіре алмады.
Жасушаларды өсіру принципін алғаш рет 1902 жылы Габерланд сипаттады. Ол жапырақтың паренхималы жасушаларын өсірді. Қоректік орта ретінде Кноп ерітіндісіне сахароза, аспарагин және пептон қосып қолданды.
Өсімдік мүшелерін және жекелеген бөліктерін өсіруде ботаниктердің қолданған алғашқы коректік ортасы өте жабайы болып табылды. Өйткені оның негізгі құрамы минералды тұздардың қоспасы болып табылатын. Өсімдік ұлпаларын бір мезгілде өсіруде В.Кноп Германияда және В.Роббинсон Америкада 1922 жылы айтарлықтай жетістікке жетті.
Бұл тақырыпты зерттеуде елеулі үлестерін қосып жүрген ғалымдар Deurenberg J.J., Zenkteler M., Melchers G., Pfahler PL., Linskens HF., Schoot HW., Wilcox M., Overbeeke N., de Waard JH., Kool AJ., de Haas JM., Hille J., Kors F., van der Meer B., Kool AJ., Folkerts O., Nijkamp HJ., Britsch L., Ruhnau- Brich B., Forkman G., J Suwinska A., Lenartowski R., Smolinski DJ., Lenartowska M.
Жоғары сәндік көрсеткіштерге сай отырғызылатын материалдарды сапалы өндіру мәселелерін шешу жолдарының бірі биотехнологияның әдістері болып табылады. Биотехнологиялық әдістер клондау, күрделі көбейетін сәндік өсімдіктердің түрлерінің санын жылдам арттыруға мүмкіндік береді.
Ғылыми жұмыстың мақсаты: Арқалық қаласында Petunia Juss. туысының түрлерін in vitro әдісін қолдана отырып санын көбейту және қаланы жаппай көгалдандыруға үлес қосу болып табылады. Алдымызға қойылған мақсатқа жету үшін төмендегі міндеттерді шешуіміз керек:
- әлемде петуния түрлерін көбейтудің заманауи әдістерін үйрену;
- өсімдік материалын дайындап, эксплаптты бөліп алу;
- эксплантты агары бар ортаға отырғызу және өсіру;
- асептикалық өскіндерден каллус ұлпаларын алу;
- өсімдіктерді микроклонды көбейту және отырғызу;
- Арқалық қаласының климаттық жағдайына төзімділігін, экологиялық факторлардың in vitro әдісімен өсірілген Petunia Juss. туысы түрлеріне әсерін зерттеу;
- Арқалық қаласын безендіру мақсатында in vitro әдісімен петуния туысы түрлерін жылдам көбейту;
Жұмыстың зерттеу әдістері:
- Теориялық (зерттеу мәселесі бойынша биологиялық, философиялық, психологиялық, педагогикалық әдебиеттерді салыстырмалы талдау, зерттеу нәтижелерін теориялық талдап қорытындылау және нәтижені жобалау);
- Эмперикалық
- жоғары оқу орнындағы озық іс-тәжірибелерді зерделеу, диагностикалау және талдау;
- тәжірибелік-эксперименттік жұмыс жүргізу;
- статистикалық (зерттеу бойынша алынған нәтижелерді математикалық- статистикалық тұрғыдан өңдеу);
Жұмыстың жаңалығы: Алғашқы рет Арқалық қаласында Petunia туысы түрлерін in vitro әдісімен көбейту жолдарының мүмкіндіктері зерттелінеді.
Жұмыстың практикалық маңызы: Жұмыстың нәтижесінде Petunia туысы түрлерінің сапалы, Арқалық қаласының климат жағдайына бейімделген көшеттерінің санын арттырып, жаппай безендіруге мүмкіндік тудырады.
І. ӘДЕБИЕТКЕ ШОЛУ
Petunia Juss. (Петуния) - алқалар тұқымдасына жататын көп не бір жылдық шөптесін немесе бұта тәрізді аспалы өсімдіктер туысы. Өсімдіктің петуния (порт. petúnia) атауы Бразилияның тупи-гуарани тіліндегі «petun», яғни «темекі, шылым» тіркесінен шыққан. 35 түрі белгілі. Биіктігі 20-30 см, кейде 60-70 см келеді. Бұл өсімдіктер жылу және жарық сүйгіш өсімдіктер. Petunia өсімдігі күтім талғамайды және ұзақ гүлдеп тұрады.
-
Клеткаларды іn vіtro жағдайында өсірудің жалпы сипаттамасы
Жаңа өсімдік материалынан бірінші рет каллус алу өте қиын, себебі әр түрлі өсімдіктердің ұлпалары, тіпті бір өсімдіктің әр бөлігінен алынған ұлпаларының дедиференциалануы және каллусогенезі жүру үшін, әр түрлі қоректік орталарды талап етеді, әсіресе ондай талап гормондардың мөлшері мен ара қатынасына қойылады. Кейбір өсімдіктердің, ұлпаларын кайталап отырғызып ұзак, мерзімде өсіргенде, ондай ұлпалар әжептәуір мөлшерде ауксин мен қатар цитокининді синтездеу қабілетіне ие болады, яғни бұл гормондарға байланысты автотрофтарға айналады. Бұндай каллус клеткалары "қалыптасқан",-деп аталады. Гормондарға тәуелсіз штаммдардың пайда болу себебі өлі белгісіз. Олардың шығуы мутациялардың әсерінен, немесе жаңа гендердің іске қосылуынан болуы мүмкін. Ісік клеткаларын жасанды ортада өсіргенде, оларда гормондарға тәуелсіз.
Клеткаларды қоректік ортада өсіргенде каллустық клеткаларға айналады, бірақ ондай жағдайда барлық клеткалар біркелкі болмайды. Жасанды ортада өскен клеткалардың түрлілігі/гетерогенді/ морфология, физиология және генетика түрғысынан болуы мүмкін.
Өсу процессін түрлі факторлардың қатысуымен реттеуге болады және мынадай көрсеткіштер арқылы бақылауға болады: өлшем, көлем, масса, клеткалар саны, белоктың және ДНҚ-ның мөлшері. Тәжірибенің мақсатына байланысты, келтірілген көрсеткіштердін, біреуі қолданылады. Клеткалардың өсуін зерттеп білу үшін, белок пен ДНҚ мелшерін белгілі бір әдістермен анықтаған ыңғайлы және оны клеткалар санын анықтағанда да қолдануға болады.
Өзіне тән бір қалыпты өсуімен және көбеюімен сипатталынатын ретсіз өсетін каллус клеткалары қайта дифференциялану процестері арқасында, ұлпалар (гистогенез), мүшелер (органогенез) және ұрық тәріздес құрылымдар-эмбриондар (эмбриогенез, немесе сомалық эмбриогенез) түзе алады.
Іn vitro жағдайында өсіретін клеткалар өсімдіктер биотехнологиясында екі бағытта қолданылады: біріншісі- клеткалар массасы, яғни каллус; екіншісі- регенерант өсімдіктері.
in vitro көбейтудің этаптары микрокөбейтудің барлық бастапқы
өсімдіктерді клондау арқылы процесстерін 4 кезеңге бөлуге болады:
Бірінші этап. Өсімдіктердің бастапқы ұлпаларының экспланттарын іn vіtro өсіру. Бұл кезеңде қоректік ортада инфекциядан таза ұлпаларды өсіріп, олардың тіршілігін сақтап, экспланттардың тез өсуіне қол жеткізу керек. Өсімдіктерді көбейтуде жетістікке жету, эксплантты дұрыс тандап алудан басталады, бұл кезде донорлық өсімдіктің өсу фазасы және өсу жағдайларын ескеру керек.
Екінші этап. Нактылы микрокөбейту кезеңі, яғни эксплантта бастама клеткалар /инициальдар/ санын көбейтіп, олардан өркендердің пайда болуына жағдай жасау.
Үшінші этап. Көбейтілген өркендерді тамырландыру және оларды сақтау. Бұл кезенде тамыр жүйесінің қалыпты өсуіне толык жағдай жасалады, қоректік ортаға тамыр пайда болуына жауапты фактор-ауксин косылады. Одан кейін өсімдіктерді топыраққа отырғызуға дайындау басталады немесе сақтау үшін төмен температура жағдайына ауыстырады. Каллусты алу жəне оны өсіру.
Кез келген өсімдіктің тірі ұлпасын қолайлы /лайықты қоректік заттар мен гормондары бар/ ортаға отырғызса, одан маманданбаған клеткалар колониясы пайда болады, оны каллус деп атайды. Каллус-қоректік ортадағы клеткалардың ретсіз бөлінуінің нәтижесінде пайда болған ұлпа. Бұрыннан бар клеткалардың көбейуінен жаңадан клеткалар мен ұлпалар тузілуін пролиферация деп атайды. Сөйтіп каллус пролиферацил нәтижесінде шығады.
Жасанды қоректік орта жағдайында, яғни іn vitro жағдайында өсімдіктердің барлық ұлпалар типтері клеткалары каллус клеткаларына айналады. Бұл процесс дифференциялану деп аталады, оның есебі фитогормондарға байланысты. Қөректік ортаның құрамын өзгерте отырып, ретсіз өсіп жатқан ұлпалардан регенерант- өсімдік алуға болады. Сонымен бір клеткадан тұтас бір өсімдікті өсіріп алуға болады, яғни өсімдік клеткалары ерекше касиетін - тотипотенттілігін көрсетеді.
Каллус ұпаларының әр типті морфогенезге икемді болғандықтан оларды көптеп өсіреді. Каллустың түзілуі мен өсуін ауксиндер мен цитокиниңдер реттеп отырады. Көптеген жағдайда каллус түзілуі үшін кейінгі өсуге қарағанда ауксиндерді 10 есе артық қажет етеді. Каллус клеткалары өсу қарқындылығымен ерекшеленеді, түсі арқылы /ақ- ақшылдан -қара қоңырға дейін, тығыздығына қарай /бос- борпылдақ, тығыз, т.б. түрлері жарықта түсін өзгертіп көгеретіндігімен тағы сол секілді белгілерімен бір-бірінен ажыратылады. Каллус ұлпаларды 3-4 апта өткеннен кейін, коректік ортадан шығарып алады да, бірнеше бөлікке бөліп, басқа жаңадан дайындалған қоректік ортаға отырғызылады, себебі қоректік ортаның күші әлсірейді, ұлпаның қоректенуі және ұлпаның ішкі бөліктерінің аэрациялануы төмендейді. Жаңа қоректік ортаның құрамы эксперименттің мақсатына байланысты болады, Егер каллустың бір қалыпты өсуін сақтап отыру керек болса, онда бастапқы қоректік орта құрамы өзгермейді.
Кейбір тәжірибелерде клеткалардың бір-біріне тигізетін әсерін ескере отырып, каллустарды өсірудің арнайы әдістері қолданылады. Қоректік ортада өсіру әдісінің бірі " бағушы" немесе " асыраушы қабат", яғни бір каллус өсу үшін өсіру әдісінің бірі - екінші бір каллус ынталандырушы ретінде қолданылады. Бұндай жағдайда ұлпаны қоректік орта сіңген фильтр қағазына салады да, өсімдіктің сол түрінің жаксы өсіп тұрған каллус массасына отырғызады. Кейде бір ыдыстың ішінде әр түрлі өсімдіктердің ұлпаларын өсіруге болады. Кейбір жағдайларда ұлпаларды бір-біріне немесе мүшелерді ұлпаларға телуді қолданады. Жақсы өсіп тұрған ұлпалар мен мүшелерден бөлініп шыққан кейбір заттар, тәжірибеге алынған каллустың өсуін жылдамдатады.
Өсімдіктің өсуі клеткалардың, ұлпалардың және мүшелердің өсуінен тұрады. Солай бола тұрса да, қандай да болсын өсудің негізі клеткадан басталады. Клетканың өсуі бірінен соң бірі кезекпен келіп тұратын процесстерден тұрады: клетканың бөлінуі, протоплазманың өсуі, созылуы және дифференциялануы. Өсу процестері негізінен меристемада жүреді.
Клетка бөлінгеннен кейін, әрбір жас клетканың алдында үш мүмкіндік болады. 1. Эмбриондық жағдайда қалып, клетка цикліне түсіп, митоз арқылы қайта бөліне алады. 2. Циклден тыс қалып, /О0/бөлінбей, тыныштық кезеңіне өтеді. З. Компетенцияға иеленіп жаңа бағыты біртіндеп айқындалып одан кейін ерекшелену /дифференциялану/ жолына түседі.
Компетенция- клеткалардың, ұлпалардың, мүшелердің, организмнің индукторлық әсерді қабылдап алуға кабілеттігі және әсер еткен факторға жауап ретінде өзінің даму бағытын өзгертуі.
Өзгерістерді әр түрлі факторлар туғызуы мүмкін. Мысалы: гормондар, көрші клеткалардың немесе басқа ұлпалардың, метаболиттері, электрофизиологиялық сигналдар және т.б.
Клетканың белгілі бір тұқым куалауды жүзеге асыруға әзір тұру қалпына тусуі-детерминация деп аталады. Қабілеттілігі бар клеткалық детерминациялануы оның дамуының белгілі бір жолын калыптастырады да, жетілудің басқа бағыттарда жүруін тежейді. Қабілетті клетканың детерминациялануы клетка бөлшегінен кейін бірден басталуы мүмкін яғни протоплазманың өсуі алдында. Белгілі бір ретпен детерминацияланған клетка өте бір тар мамандыққа ие болады яғни дифференцияланады (қандай да бір ұлпаның клеткаларына айналады).
Дифференциация - жас клеткалар арасындағы құрылысы мен кызметі жағынан болатын өзгерістерді,аналық клеткалары мен жаңа пайда болған клеткалар арасында айырмашылықтарды тудыратын күрделі процесс. Меристемалық клеткалар атқаратын қызметі және құрылысы жағынан біркелкі, дифференциялану нәтижесінде әр жолдармен дами бастайды да,түрлі мүшелердің ұлпаларын түзеді. ДНҚ-ы молекуласында шифрленген генетикалық информацияның іске асуы арнайы и-РНҚ түзілуі /транскрипция/ арқылы және одан кейін генетикалық кодына сәйкес белоктар синтезі /трансляция/ арқылы жүретіні белгілі. Бірақ, барлық генетикалық информация бір уақытта іске аспайды, клетканың өсу және дифференциялану процесстері кезінде қатаң тәртіппен кезектесіп көрініп отырады. Геномдағы гендердің бір тобы бір кезеңде активті болады, яғни иРНҚ-ның мағыналы өнімі бар, ал келесі бір тобы активтілігін көрсете алмайды, себебі, ондай гендер активтілігі репрессор әсерінен тежелген. Сондықтан, клеткалар арасындағы айырмашылықтар, яғни олардың дифференциаялануы гендердің әр түрлі дифференциалды/ активтенуіне байланысты.
Морфогенез дегеніміз пішін /форма/ құру процесі, онда өсімдік мүшелерінің бастапқы элементтерінің құрылуы, өсуі және өнуі /органогенез/, ұлпалардың /гистогенез/ жөне өсімдік клеткаларының жетілуі /цитогенез/ немесе клетканың дифференциялануы жүреді. Морфогенез көп кезеңдерден тұратын күрделі процесс.
Организмнің дамуы - өсу, дифференциалануы, морфогенез процесстерінің бір - бірімен тығыз байланыста жүруінің нәтижесінде болады. Бүтін организмде бұл процесстерді бір - бірінен ажыратып қарау өте қиын. Сонымен қатар, бұл процесстер бір - біріне мүлде ұқсамайды, әрқайсысының өзіндік ерекшеліктері бар. Атап айтқанда сандық өзгеріс және сапалық жағынан қайта құрылудың нәтижесінде кеңістікте құрылысы және формасы бар құрылым пайда болады, онда морфогенез процессі жүреді.
К. Уоддингонның анықтамасы бойынша, "Морфогенез дегеніміз - гомегендік, яғни жеке бөліктерден тұрмайтын массасының белгілі бір құрылымға айналуы". Тұтас организмде әр түрлі бөліктердің бір - біріне тигізетін әсерін камтамасыз ететін, көптеген корреляциялар, байланыстар болғандықтан, бұндай жүйеде морфогенездің көрінуін зерттеу өте қиын шаруа.
Морфогенезді зерттеудің өте қолайлы моделі, ол ұлпаларды, клеткаларды және протопласттарды жасанды қоректік ортада өсіру. Моделді қолданып жүргізген зерттеулердің артықшылығын, ол өсірілетін клеткаларға бір бағытта әсер ету арқылы, зерттеуші түрлі факторлардың әсерін бақылай алады.
Каллустың пайда болуына əсер ететін факторлар
Жасушалардың өсуіне бірнеше физика- химиялық факторлар әсер етеді. Физикалық факторларға: температура, қысым, араластыратын айналу жиілігі, көбік түзілу, ау қабығының жылдамдығы, қоректік ортаны, субстратты беру жылдамдығы, тұтқырлығы; Химиялық факторларға: қоректік ортасының құрамы және рН көрсеткіштері, тотығу- тотықсыздану потенциалы, еріген оттегімен көмірқышқыл газының мөлшері, көміртегі, азот, фосфор, магний, калий, кальций, натрий, темір, т.б иондармен тұздар құнарлығы жатады. Қазіргі микробиологиялық өндірісте ең жиі қолданылатын реактор субстрат пен жасушалар араласатын резервуар болып табылады, онда реакциялар жүру үшін қолайлы жағдайлар жасалынады, температура мен рН көрсеткіштері реттеледі. Қоректік орта арқылы кейде оттегімен қаныққан, сүзілген ауа айдалады, химиялық және биологиялық талдау үшін екі тәсілі: 1. барлық кіретін саңылауларды залалсыздандыру. 2. реактор ішінде атмосфералық қысымнан артық қысым жасалыну қолданылады. Бірнеше сағаттан бірнеше күнге дейін созылатын үдеріс аяқталғанда, барлық қоспаны реактордан алып тастайды, өнімді бөліп, тазалауды жүзеге асырады (1- кесте).
1- кесте. Петуня көшетінің тауарлы өнімділігі мен сәндік бағасына қоректік орта мен себу тәсілінің әсері
Қоректік орта
Себу тәсілі
Өсім-діктің тығыз-дығы, дана/м2
Тауарлы өнім, дана/м2
Сәндік бағасы
1
Аммоный селитрасы-10 г/м2+ қос суперфосфат -20г/м2,+ хлорлы калий- 15г/м2
Шашып себу
1250
625
Нашар, сүйкімсіз
Пунктирлі себу
870
609
жақсы
Пластмас ыдысқа (стаканға) себу
650
595
Жақсы, тартымды
2
Аммоный селитрасы-10 г/м2+ қос суперфосфат -20г/м2,+ хлорлы калий- 15г/м2+ бор қышқылы- 0,2г/м2
Шашып себу
1230
639
Қанағаттанарлық
Пунктирлі себу
860
633
Жақсы
Пластмас ыдысқа (стаканға) себу
650
598
Жақсы, тартымды
3
Аммоный селитрасы-10 г/м2+ қос суперфосфат -20г/м2,+ хлорлы калий- 15г/м2+ бор қышқылы- 0,2г/м2
Шашып себу
1240
630
Қанағаттанарлық
Пунктирлі себу
860
645
Жақсы
Пластмас ыдысқа (стаканға) себу
640
615
Жақсы, тартымды
Оқшауланған жасушалар, ұлпалар, мүшелер өсуіне қоректік ортадан басқа да жағдайлар әсер етеді. Өсімдік жасушаларын қатты және сұйық ортада өсіру үшін, оның рН көрсеткішінің маңызы өте зор. Табиғи жағдайда жасушаның тіршілік әрекеттері сутегі иондарының қолайлы концентрациясында (рН 5,5- 7,5) өтеді. Қоректік орталардың буферлік сиымдылығы өте төмен, сондықтан олардың алғашқы рН көрсеткіші өсімдік жасушаларында өсіргенде тез бірнеше бірлікке жылжып кетеді. Соның арқасында жасушаның биосинтездік қасиеттері өнімдердің жиналуы өзгереді. Сонымен қатар, агарланған қатты қоректік ортаны дайындағанда агар қышқылы ортада ғана қатады, яғни агардың полимеризациясы рН 5-6 сәтті өтеді.
Өсімдік жасушаларын өсіру үшін 25 0С температура қажет. Жасушалардың өсуіне әсер ететін сыртқы факторлардың бірі- жарық. Жалпы in vitro жағдайында өсірілетін өсімдік жасушаларында жасыл пигмент хлорофилл түзілмейді, сондықтан олар әдеттегідей фототрафтық жолмен емес гетеротрофты қоректенеді. Жасушаларда қосымша заттар түзілуіне жарықтың сапасы, қарқындылығы және фотопериодтың әсер ететіндігі дәлелденген, сондықтан жасушалық технологияны жасаудағы басты мақсаттардың бірі- жасушаларды өсіру үшін қажетті жарықтың сапасы мен қарқындылығын анықтау.
Жасушалардың өсуіне аэрацияның әсері зор (бұл тұрғыда арнайы суспензияға қажет). Аэрация болмаса, жалпы алғанда, суспензияның өсуі мүмкін емес. Клеткалар суспензиясы (лат. Suspension- асып қою)- жеке жасушаларды немесе кішігірім жасушалар тортарын аппаратура арқылы ауамен қамтамассыз етіп және араластыра отырып сұйық қоректік ортада өсіру. Жасушаларды өсіргенде қоректік ортаның осмос қысымын да ескеру керек. Жоғары осмос қысымы қоректік заттарды жасушалардың сіңіруін қиындатады. Экспланттан пайда болған морфогенді каллус ұлпаларды әрбір 3-4 апта сайын жаңа қоректік ортаға көшіріп отырса, олар шексіз ұзақ өсе береді. Осындай морфогенді каллусты жарықта, 27 0С температурада өсіргенде регенерант- өсімдік дами бастайды.
Petunia биіктігі 20-30 см, кейде 60-70 см келеді. Бұл өсімдіктер жылу және жарық сүйгіш өсімдіктер. Өсу температурасы 18- 26 0С аралығында. Яғни, Арқалық қаласының климаттық жағдайы Petunia өсімдігін өсіруге қолайлы болып табылады. Арқалық қаласының жылдық жауын-шашын мөлшері 270 мм, қаңтарда орташа температура 17 С градус, шілдеде 21-24 С градус (2- кесте).
2- кесте. Гүлдеу уақыты мен сәндік бағасына тұқым себу мерзімі мен температураның әсеріТұқым себу мерзімі
Өсіру температурасы
Гүлдеу уақыты
Сәндік бағасы
1
Ақпанның бірінші он күні
16-180С
Сәуірдің бірінші он күндігінде
Қанағаттанарлық
20-220С
Сәуірдің бірінші он күндігінде
Қанағаттанарлық
24-260С
Сәуірдің екінші он күндігінде
Қанағаттанарлық
2
Ақпанның екінші он күні
16-180С
Сәуірдің екінші он күндігінде
Қанағаттанарлық
20-220С
Сәуірдің екінші он күндігінде
Жақсы
24-260С
Сәуірдің екінші он күндігінде
Қанағаттанарлық
3
Ақпанның үшінші он күні
16-180С
Сәуірдің үшінші он күндігінде
Қанағаттанарлық
20-220С
Мамырдың бірінші он күндігінде
Жақсы
24-260С
Мамырдың екінші он күндігінде
Қанағаттанарлық
ІІ. ЗЕРТТЕУ ӘДІСТЕРІ
2.1. Өсімдіктер биотехнологиясында залалсыздандыру жұмыстарын жүргізу әдістері
Оқшауланған жасушаларды, ұлпаларды, мүшелерді және протопласттарды өсіруде нақты жетістіктерге жету үшін қатаң залалсыздандыру жағдайы сақталынуы тиіс. Өйткені өсімдік жасушалары мен ұлпаларын өсіруге арналған жасанды қоректік ортада микроағзалар жақсы дамиды да нәтижесінде өсірілетін материалға зиян келтіреді.
Микроорганизмдердің тіршілік әрекетінің нәтижесінде қоректік ортаның құрамы өзгереді. Сонымен қатар өсірілетін жасушалар мен ұлпалар, протопласттар микроорганизмдермен бүлінеді.
Сондықтан өсімдіктің оқшауланған жасушаларын, ұлпаларын, мүшелерін және протопластарын өсіруге арналған барлық тәжірибелерді залалсыздандырылған ламинар-бокстарда жүргізеді. Боксты, саймандарды, ыдыстарды, өсімдік материалдарын, қоректік ортаны, мақталы тығындарды және басқа да жұмыс жасауға қажетті материалдарды мұқият залалсыздандырады (3- кесте).
3- кесте. Өсімдіктерді залалсыздандыру ерекшеліктері
Залалсыздандырғыш заттар
Диоцид-0,1%
Сулема-0,1 %
N8 гипохлориті
5-9 %
Сутегі
пероксиді 12 %
Құрғақ ұрықтар
15-20
10-15
15-20
12-15
Ісінген үрықтар
6-10
6-8
10-15
6-8
Түбірлі жемістің етті ұлпалары
20-30
15-20
15-20
-
Сабақ үлпаралы
20-40
20-25
2025
-
Жапырақтар
1-3
1-3
3-6
3-5
Апекстер
1-10
1-7
3-15
2-7
Ламинар - боксты залалсыздандыру. Алдымен ламинар бокс ішіндегі жұмыс орнын 70% спиртпен сүртіп шығамыз. Ламинар боксқа спирттік жандырғышты, шырпыны, 96 процентті спирті бар стаканды және залалсызданған суы бар колбаны апарып қоямыз. Меристеманы алу үшін бинокулярлы үлкейткіш шыныны дайындайды. Жұмыс жүргізер алдын 10-12 сағат бұрын ламинар бокстың ішін бактерицидті ультракүлгінді сәулелі лампамен өңдейміз.
Ыдыстарды залалсыздандыру кептіргіш шкафта немесе автоклавта жүргізіледі. Залалсыздандырмас бұрын ыдысты жуып кептіріп аламыз. Ыдыстарды жуу үшін детергенттер және хромпик қолданылады. Пробиркаларды колбаларды залалсыздандыру алдында мақта тығындарымен аузы жабылады да орама қағазбен оралады. Содан соң ыдысты кептіргіш шкафқа салып +175°С температурада 2 сағат бойы қыздырамыз. Бұл температурада бактериялармен қатар спораларда жойылады. Кептіргіш шкафтың температурасын 175°С жоғарылатудың қажеті жоқ, өйткені тым жоғары температурада мақта тығын сарғайып, орама қағаз сынғыш болып қалады.
Автоклавтағы жоғары кысымының нәтижесінде бұдан да жоғары сапалы залалсыздандыруға болады, себебі ылғалды ыстықта микроорганизмдер мен споралар тиімді жойылады. Автоклавта тығыны бар стакандарды, Петри табакшасын, пипеткаларды, суы бар колбаларды залалсыздандыруға болады. Ыдыстарды орама қағазға немесе фольгаға орап 2 атмосфералық кысымда 25-30минут бойы залалсыздандырады. Пипеткалардың ашық жағын мақта тығынымен жауып, қағазға 10 данадан орап залалсыздандыру керек.
Құралдарды залалсыздандыру. Құралдарды (скальпельдерді, пинцеттерді, инелерді) алдын-ала залалсыздандыру 140-160°С 2 сағат бойында құрғатқан шкафқа құрғақ ыстық бумен ысытып алады. Шприцтер мен қайшыларды, тескіштерді (еlеrlа) қайнатқан жөн. Металды заттарды автоклавирлеуге болмайды: өйткені олар бу әсерінен тот басып өтпей қалады. Жұмыс алдында және жұмыс барысында құралдарды (пинцеттерді, скальпельді, микробиологиялық инелерді) тағы да бір залалсыздап алу керек. Залалсыздауды спирт жалынмен күйдіре отырып, 96 процент этил спирті бар фарфорлы стканы бар ламинар-бокста жүргізеді. Бұдан кейін әр бір құралдарды алдын-ала залалсыздандырылып дайындалған қалың қағаз арасында орналастырады. Залалсызданған құралдарды тек бір ғана рет пайдаланады. Екіншілей қолданар алдын оны қайта спиртпен залалсыздандырып күйдіріп алады. Өте жұқа құралдар (ине, алмастың бір түйірін) күйдіру барысында өз қасиеттерін жоғалтып алуы мүмкін, сондықтан оларды спиртке батыру аркылы залалсыздандырады.
Материалдар ұлпаларды жасушаларды оқшаулау кезінде пайдаланылатын мақта, марле, мақта тығындары, сүзгіш қағазы, халаттар, қалпақтар автоклавта 2 атмосфералық қысымда 25-30минут бойы залалсыздандырылады.
2.2. Клеткалардың қоректенуі және қоректік орталардың жалпы сипаттамасы
Өсімдік клеткаларын өсіруге арналған қоректік ортаның құрамы өте күрделі. Ол құрамға минералды тұздар, көмірсулар, амин қышқылдары, витаминдер, түрлі өсуді реттегіштер кіреді. Әр өсімдік ұлпасы өсуі, сонымен қатар бір өсімдіктің, түрлі мүшелері және ұлпаларының өсуі үшін де қоректік ортаның, белгілі бір құрамын талап етеді. Пайда болған каллусты одан әрі өсіру үшін, сонымен қатар органогенез процессін қоздыру үшін де, қоректік ортаның құрамын өзгертіп отыру қажет. Зерттеуші өз жұмысы барысында зерттелетін обьектіге сәйкес қоректік ортаны жетілдіруге талаптанады, сол себептен қоректік ортаның саны да, түрлері де көп (4- кесте)
Қоректік ортаның құрамына минералды тұздар макро және микроэлементтер түріде кіреді. Көптеген қоректік орталар құрамына азотнитрат немесе аммони тұздары, фосфор-фосфат, күкірт сульфат түрлерінде, ал темір анорганикалық тұздар және хелат түрінде қосылады. Барлық қоректік орталар құрамында сонымен қатар К+, Са2+ және Ме2+ иондары болады, бірқатар микроэлементтер кіреді. Микроэлеметтер әсіресе сұйық қоректік ортада өсетін суспензиялық клеткаларға аса қажет.
Ұлпалар мен өсімдік жасушаларын өсіру орталарының компоненттерін (жинақтарын) 6 негізгі топқа бөлуге болады, бұл әдетте макроэлементтерді, микроэлементттерді, темір көзін, витаминдерді, көміртегі көздері мен фитогармондардың концентрлі ерітінділерін дайындау ретіне байланысты болады.
Өсімдік экспланттарын культивирлеу үшін барлық қоректік орталарға минералды тұздар қоспалары негіз болып табылады.Бұл азот қоспасының нитрат, амони тұздары, нитрит түрінде,фосфор-сульфат түрінде; сондай-ақ ертілетін тұздар K+,Na+, Ca++, Mg ++ -болып келеді.Темір хелат түрінде [FeO4 немесе Fe2O4 +ЭДТА (этилендиаминтетрасірке қышқылы) қолданылады немесе ас тұзы NaЭДТА(трилон-Б)] өсетін ұлпалармен игеріліп кетуіне мүмкіндік береді.
Азот, фосфор, күкірт органикалық қосылыстардың яғни ақуыз, май, нуклейн қышқылдар құрамына жатады. Темір, цинк, марганец, молибден, кобальт порфириндер қасында тотығу-тотықсыздану ферментттері (каталаздар, периоксиездар, полифенолоксидаздар) фотосинтез макромолекулаларын құрайды.Осыған орай,барлық қосылыстар жасушалар мен ұлпаларда структуралы функциялы орындайды.Сонымен бірге K+, Na+, Ca++, Cl-, H+ иондары ортаның рН реттеуге және жасушалардың физиологиялық градиенттерін (тургорды, полярлықтың осмотикалық қысымын) ұстап тұру үшін қажет.
Биологиялық макромолекула үшін, сондай-ақ гетеротрофты ұлпаларды культивирлеудегі көміртегі көзі ретінде (каллустың және суспензияның) қоректік ортаға 20-60г/л концентрлі көміртектерін қосады. Әдетте бұл дисахаридтер (сахароза), моносахаридтер (гексоздар: глюкоза және фруктоза, кентонздар: ксилоз бен және т.б.). Полисахаридтерді қоректік орталарда қолдана бермейді.Тек гидролитті ферменттерді құрайтын кейбір ұлпалар түрін (ісікті) крахмалмен, целлобиозы бар орталарда ғана өсіреді.
Биохимиялық реакцияларға жағдай жасау үшін жасушада витаминдер тобын (В1,В6,В12) С (аскорбин қышқылы) РР(никотин қышқылы), мезоннозиттер сияқты биологиялық катализаторды қолданады.
Тиамин (В1) пируватексрбоксилаз құрамына кіреді және көмірсуларына айналу барысына қатысады.Тиаминпирофосфат кетоқышқылының тотықтырғыш декарбоксилирлену ферменттер құрамына кіреді және транскетомездар кофертенті болып табылады.
Пиродоксин (В6) фосфорқышқылды эфир түрінде ферменттің декарбоксилирлеуі мен аминқышқылдарының аса аминирлену құрамына кіреді.
Никотин қышқылы (РР) амид түрінде катализденетін донорлы-акцепторлы тізбе Н+(органикалық заттар молекуласынан Н+алу), НАД және НАДФ дегидрогенез құрамына кіреді.
Ұлпалар культурасындағы түр түзуші процестерді дамыту үшін биологиялық даму реттегіштері мен фитогармондардың жетілулері қажет. Бұл заттар жасушалар мен ұлпалардың дедифенциясына және дифференциясына әсерін тигізеді,гистогенезді қорғайды, жасушалардың созылуын және жіктелуді индуцирлейді, жетілу мен қартаю процестеріне қатысады, немесе жағдай жасайды болмаса дамушылық қабілетін және жасушалы культураның жетілуін ингибирлейді, жыныстық қалыптасуды шарттастырады. Биотехнологиялық ізденістерде көбінесе ауксиндер, цитокининдер және гиббирилиндердің дамуы мен жетілуіне жағдай жасайтын гармондарды пайдаланады.
Ауксиндер: ИУК-β индолил-3-сірке қышқылы, ИМК-индолил-3-май қышқылы, НУК-α-нафтилсірке қышқылы, 2,4-Д-2,4-дихлорфеноксисірке қышқылы.
Цитокининдер кинетин-6-фурфуриламинопурин, зеатин, NH-дифенилмочевина, 6-БАП-6-бензиламинопурин, 2-изопентениладенин(2 ір).
Гибберилиндер: гибберелді қышқыл.
Алғашқы каллус индукциясы үшін қажетті биологиялық қоспа ретінде (ортаның жалпы көлемінен 10-15%) өсімдік экстрактын; кокос сүтін (кокос жаңғағының сұйық эндоспермі) зеатин және кинеин цитокининдерді құрайтын пісіп жетілмеген жүгерінің дәнінен шығатын (ең жақсы сүт кезінде) ( орнын басатын 6 аминопурининдер) және N-дифенилмочевинаны қолдануға болады. In vitro культурасында сұйық және қатты орталарды қолданады. Сұйық орталар суспензияларды, каллусты, оқшауланған денелер мен ұлпаларды регенерант өсімдіктерін культивирлеу үшін пайдаланады. Бұл үшін пробиркалаларға синтетикалық сілтілеу материалдарды немесе сүзгіш қағаздардан арнайы ұстап тұрғыш орналасады.
Қатты (агарланған) ортаны 5,6-6-рН-ында сумен гелді құрайтын теңіз балдырлары құрамына кіретін агар-агармен-полисахарид негізінде дайындайды, ал кейде тығыздығын және орын алмастырғыш ретінде Р10,Р100 (биогель) полиакриамильді гельдерін қолданады.
Жасанды қоректік орта үшін қажетті макро және микро тұздар алдын-ала дайындайды және бірнеше рет пайдаланады. Бұл концентрлі ертінділер. Оларды арнайы орындарда: макро және микро тұздарын 0+40С мұздатқышта тесік жабығы бар ыдыста сақтайды, витаминдерді, фитогармондарды, ферментттерді өсінді экстракталарын 200С-та жабыны бар 5-10 мл ыдыстарда сақтайды (пеницилин флокондары).
Макро тұздардың концентрлі ертінділері. Әдетте жұмнатқарушы концентрациясы бойынша 10-40 есеге, микротұздар 100-1000 есеге,витаминдер -1000 есеге асып түседі.
Фитогармондар ертінділерін қоректік орталармен бірге жұмыс алдында дайындаған дұрыс болады.
Макро және микро тұздардың концентрлі ертінділерін дайындау үшін әрбір тұздарын және стаканға салып ысыту барысында ерітеді, одан соң құйып алып қажетті бір белгіге дейін жеткізеді.Суытылған микротұздар қоспасына молибден тұзының ертіндісін қосады (тұнба түзуіне мүмкіндік бермеу үшін).
Темір хелатының және кальций хлоридінің концентрлі ертінділерін (күкірт қышқылды темір+ЭДТА, немесе ЭДТА-трилон-Б)басқа тұздардан бөлек сақтап дайындайды.
Витаминдердің концентрлі ертінділерін келесідей тәртіппен дайындайды:10 еселі өлшенген затты 10 мл дистильденген суда әрқайсысын бөлек-бөлек ерітіп алады.
Фитогармондар- бұл суда нашар еритін зат. Сондықтан 100мг затты алдын-ала ащдаған мөлшердегі (0,5-1,2 мл) спиртте (ауксиндер мен гибберилиндер), 0,5н HCl немесе KOH (цитокининдер), одан соң (абезинді қышқыл мен кинетининнен баққаны) толық ерігенге дейін қыздырады және көлемін 100 мл көлемге дейін жеткізеді (1мл-1мг затты құрайды).
Жасушаларды, ұлпаларды және өсімдік денелерін культивирлеу үшін әр түрлі құрамдағы қоректік орталар қолданылады. Көбінесе Мурасиге-Скуга, Уейт, Гамборг (В5) орталары кеңінен қолданылады.
Ұлпалар жасанды қоректік ортада гетеротрофтар болғандықтан, оларға қоректену үшін көмірсулар кажет. Көмірсулардың және энергияның көзі ретінде негізінен концентрациясы 20-40 г/л сахароза, кейде глюкоза қолданылады. Өсімдіктердің кейбір түрлері жасанды қоректік ортада қанттардың басқа түрлерін пайдаланғанмен, әдетте олардың қажетке жарауы шамалы.Көміртегі көзі ретінде бірқатар органикалық қышқылдар мен спирттер сынақтан өткізілген, бірақ олардың жарамдылығы көмірсуларға қарағанда төмен болған.
Қоректік орталардың барлығының құрамына дәрілік витамиңдер кіреді. Витаминдердің ең маңыздылары деп, В тобына жататындары есептеледі. Олардың ішінде: тиамин /В,/, рибофлавин /В^/, пиридоксин /В,/. Көптеген өсіруге алынған клеткаларға бұлардан басқа никотин кышқылы, фоли, иантотен қышқылы, мезоинозит қажет.
Көптеген ұлпалардың жасаңды қоректік ортада витаминдер синтездеуге мүмкіндік болғанмен, зат алмасу кабілетін толық көрсету үшін ол мөлшер жеткіліксіз. Сондықтан қоректік ортаға витаминдерді қосымша беру ұлпалардың өсуін ынталандырады.
Қоректік ортаның құрамы қандай күрделі болса да, оған ен қажет компоненттердің бірі фитогормоңдар, себебі бұл қосылыстар өсімдіктерде жүретін физиологиялық процесстерге қатысады.Өсімдіктер клеткалары өсуі мен дифференциялануы үшін ауксиндер мен цитокининдер керек. Өсуді реттегіштер жоқ қоректік ортада тек ісік ұлпалары және қалыптасқан ұлпалар өседі.
Қатты қоректік орта дайындау үшін, гель түзетін зат ретінде агар-агар пайдаланылады. Агар-полисахарид, құрамында көптеген қоспалары азот, минералды элементтер, витаминдер бар, сондықтан ағын судың астыңда 4-6 сағат жуу қажет. Өсіруге алынатын ұлпалардың қажетіне қарай қоректік ортадан агар-агардың мөлшері 0,8 ден 1,5 % -ке дейін болады. Ең алғаш өсімдіктердің ұлпаларын жасанды қоректік ортада өсіру агарланған қоректік ортада жақсы нәтиже берген. Бұндай табысқа 1930 жылдары француз Р.Готре және американдық ғалым Ф.Уайт ие болған. Қазіргі уақытта жер шарының кейбір лабароторияларында сәбіздің тамыр сабағынан 1938 жылы Р.Готре алған клеткалары штаммдары сақталған. Каллустың клеткаларын қанша ұзақ уақыт болса да өсіруге болады, тек оларды әрбір 3-4 аптадан кейін жаңа дайындалған қоректік ортаға кайта отырғызып отыру қажет. Осы тәріздес ұзақ, қайта алмастырылып отырылатын қоректік ортада клеткалар мен каллус ұлпалары өсіріледі.
Өсімдік клеткаларын өсірудің техникасының дамуының нәтижесінде бөлініп алынған ұлпалар мен клеткаларды суық қоректік ортада өсірудің әдістері жете зерттелді. Клеткаларды сұйық қоректік ортада өсіру немесе суспензияда өсіру деп, қалқыған клеткаларды немесе кішігірім клеткалар тобын аппаратура арқылы таза ауамен канықтырып және араластыра отырып сұйык қоректік ортада өсіруді атайды. Сұйық ортада клеткалардың таза ауамен канықтырылуының әр түрлі тәсілдері бар: ыдысты тоқтаусыз айналдыру немесе шайқады тағы бір жолы сұйық ортаға залалсыздандырылған ауа үрлеу арқылы. Клеткаларды сұйық қоректік ортада өсірудің өзіндік артықшылыктары бар: олар қоректік заттармен жақсы қамтамасыз етіледі және токсинді заттары тез бөлініп шығады. Ұлпаларды қатты немесе сұйық орталарда болсын өсіргенде, ортаның рН-ы мадызды роль атқарады, ұлпаларды өсіруде жетістікке жету үшін орта рН-ы қолайлы және тұрақты болу керек.
Макромолекулалардың активтілігі мен кұрылымы ортаның рН-на тәуелді, әсіресе ұлпаның құрамындағы ферменттік белоктарға әсері жоғары. Сонымен бірге, ортаның рН-ы қоректік ортаның компоненттерінің тұрақтылығымен оларды клеткалардың қабылдап алуына, әсіресе есуді реттегіштер мен витаминдердің сіңуіне әсер етеді. Егер ортаның рН-ы төмен болса, агар қатпайды, қоректік ортаның рН-ы белгілі деңгейге дейін, қыщқыл немесе сілті қосып жеткізіледі. Көптеген ұлпаларды өсіргенге орта рН-ы 5,5 - 5,8 тең болу керек.
Қоректік орталарды дайындағанда және клеткаларды өсіргенде кейбір физикалық факторларды ескеру керек, мысалы: қоректік ортаның осмостық қысымы, температура және жарық. Жоғары осмостық ыдысын қоректік заттардың сіңуін қиындатады. Температура метаболизм процесстеріне әсер етеді, көптеген өсімдіктерді өсіру үшін ең қолайлы температура 24-27 градус. Әдетте үлпаларды қараңғыда немесе шашыраңқы жарықта өсіреді. Тура түскен жарық өсуді бәсендетеді. Хлорофилі жоқ ұлпаларға жарықтың тигізетін әсері фитохром жүйесіне байланысты. Жасыл ұлпалардың өсуі мен морфогенезіне жарықтың тигізуіне» әсерін тек фотосинтездің активтілігімен байланысты деп түсінбеу керек, себебі, ұлпаларды қоректік ортада өсіргенде оларда жүретін фотосинтез қарқындылығы өте төмен.
Тұздардың (макро- және микроэлементтер), витаминдердің, фитогормондардың ерітінділерін дайындау әдістері
Қоректік ортаның негізгі (ингредиенттері) қоспалары ретінде макро-және микротұздар, дәрумендер, органикалық заттар, өсуді реттегіштер қолданылады. Қоректік ортаның минералдық құрамы меристеманы қажетті қоректік элементтермен толық қамтамасыз етуі қажет. Картоп эксплантының оптималды өсуіне қажетті рН мөлшері 5,7. рН мөлшері ауытқыған жағдайда 0,1н КОН немесе 0,1н НСl қосу арқылы қажетті мөлшерге келтіру қажет. Орта аздап буферлі болуы керек, себебі меристеманы культивирлеу кезінде орта кышқылданады. Өсімдіктерді іn vitro жағдайында өсіруге ең қолайлы орта Мурасиге-Скуга минералды негізіндегі қоректік орта. Одан басқа Уайт Гамборг қоректік орталары қолданылады.
Дәрумендер тобының ішінен ортаға пиридоксин В₆, тиамин В¹, аскорбин қышқылын, биотинді, мезоинозитті, пантотенат Са, никотин қышқылын (РР) қосады. Ортаның басты компонентінің (қоспасының) бірі - сахароза -меристеманы өсірудегі көмірқышқыл қорегінің көзі. Меристеманың өсуіне ортаға аминқышқылы көзін, мысалға казеин гидролизатын қосу оңды әсер етеді. Қоректік ортаға қосылған өсуді реттегіштер меристеманың өсуін айтарлықтай тездетеді. Көбіне өсуді реттегіштерді ауксиндер тобынан қосады, яғни сабақ пен тамыр ұштарының өсуін реттейтін заттар (Р -индолил-3-сірке қышқылы - ИУК) немесе ауксиннің синтетикалық аналогтары: индоломай қышқылы (ИМҚ), α - нафтилсірке қышқылы (α-НСҚ) және 2,4 дихлор феносірке қышқылы (2,4 Д). Мұнан өзге ауксиндердің өсімдіктегі физиологиялық активтілігінің концентрациясы өте төмен (0,005-100 мг/л). Өзге ауксиндер тәрізді бұлар да ризогенезге және тамырдың одан әрі өсуіне әсер етеді; әдетте 1-2 мг/л концентрациясы қолданылады. Активті көмір тамыр түзілісін реттейді - ИМҚ 0,1-1 мг/л концентрациясы қолданылады; α - НУҚ 0,5-1 мг/л концентрациясы каллус және тамыр түзілісін реттейді.
Өсімдіктердің ауксин алмасуына өсімдіктің өсуін реттейтін, лигнификацияға жағдай жасайтын фенолды қосылыстар: ферул қышқылы, кониффил спирті, кофейн қышқылы қатысады. Сабақтың өсуін гиббереллиндер реттейді. Өскінге гиббереллин сепкеннен кейін сабағына сіңеді, бірақ түйін аралық саны артпайды. Гиббереллиндер дифференцирленген ұлпаларға емес, меристемалық зонадағы жасушаның бөлінуіне әсер етеді.
Әдетте қоректік ортада - 0,2 мг/л концентрациясы қолданылады. Қоректік орталардың маңызды компонентінің бірі жасуша бөлінуін реттейтін, топырақтың дифференциациясына жағдай жасайтын, сарғайған жапырақты көгертетін, тамыр түзілісін ингибирлейтін цитокининдер -фитогормондар; қолданылатын концентрация мөлшері - 0,02-0,1 мг/л.
Қатты қоректік ортаны дайындау үшін теңіз балдырларынан пайдаланылатын полисахарид -агар-агар пайдаланылады. Қажетсіз қоспалары аз немесе мүлде болмайтын бактериальдік «Difсо» агарын алдын-ала жуусыз пайдалана беруге болады. Әдетте қатты қоректік ортаны дайындау үшін 5-8% агар қосылады. Макротұздар, микротұздар, Ғе хелаты мен дәрумендердің концентрленген ерітіндісі пайдаланылады. Алынған аналық ерітінділер тоңазытқышта, ал дәрумендер төменгі температурада сақталынады. Макротұздарды 10-20 есе, микротұздарды 100-1000 есе, дәрумендерді 1000 есе концентрленген күйінде дайындалады (4- кесте).
Кесте -4. Мурасиге және Скуга қоректік ортасын дайындау қажет .Құрамы
Мөлшері, мг/л
Құрамы
Мөлшері, мг/л
NH4NO3
1650
FeSO
27,8
KNO3
1900
Na-ЭДТФ 2H2O
37,3
CaCL2·2H2O
440
Тиамин
0,1
MgSO4·H2O
370
Пиридоксин
0,5
КН2РО4
170
Никотин қышқылы
0,5
MnSO4·4H2O
22,3
Мезо-инозит
100
CoCl2·6H2O
0,025
Глицерин
2,0
ZnSO4·7H2O
8,6
ИУҚ
2,0
CuSo4·5H2O
0,025
Кинетин
0,2
Na2MoO4·2H2O
0,25
Сахароза
30,0
Kj
0,83
Кесте- 5. Мурасиге және Скуга бойынша аналық ерітінділердің құрамыҚоректік ортаның құрамы
№
Макротұздар
Аналық ерітіндіде г/л
№
Микротұздар
Аналық ерітіндіде мг/100 мл
1
KNO3
38
1
н3ро4
620
2
СаС12 (сусыз)
8,8
2
МnS04 * 4Н20
2230
немесе СаС12 *2Н20
13,8
3
ZnS04
860
3
NН4NO 3
33
4
Кl
83
4
МgS04 (сусыз)
3,6
5
Nа2МоС14 * 2Н2С
25
немесе МgS04 *7Н20
7,4
6
СuS04 * 5Н20
2,5
5
кн2ро4
3,4
7
СоС12 * 6Н20
2,5
1л қоректік ортаға аналық ерітіндінің әрқайсысынан 50мл алу қажет
1 л қоректік ортаға аналық ерітіндінің әрқайсысынан 1 мл алу қажет
Ғе-хелат, аналық ерітіндіде г/100 мл
1
ҒеS04 * 7Н20
557
2
ЭДТА-Nа2(трилон Б)
745
1л қоректік ортаға аналық ерітіндіден 5 мл алу
Қажет
5. Фитогормондардың ерітінділерін төмендегідей етіп дайындайды;
5.1. Ауксиндер (2,4-Д, -НУК, ИУК, индолил-май қышқылы - ИМҚ): 100 мг затты 0,5-2 мл спиртте ерітеміз, аздап қыздырамыз да дистилденген сумен 100 мл дейін жеткіземіз (концентрациясы 1 мг/мл).
Цитокининдер (кинетин, зеатин, БАП): 0,5 н. НСІ аздаған көлемде ерітеміз, аздап қыздырамыз, дистилденген сумен 100 мл дейін жеткіземіз (0,1 н КОН -тың 2-3 тамшысынан артық қолданбаған дұрыс, орта тым сілтіленіп кетуі мүмкін).
5.3 Абсциз қышқылы (АБК): үлгіні 70 пайыздық этанолда ерітіп қажетті көлемге дейін жеткіземіз.
5.4. Гиббереллин қышқылы (ГҚ): үлгі суда ерітіледі.
Аналық ерітінділерді пайдаланып өсімдік мүшелері мен ұлпаларын өсіруге қажетті қоректік орта дайындалады.
Аналық ерітінділер дайындауға қажетті тұз түрлері мен аналық ерітіндінің мөлшерін 2 - кестеде көрсетілгендей етіп келтіру қажет. Дайын ерітінділерді қақпағы жабылатын шыны ыдыстарға құйып этикетка жабсырып тоңазытқышқа сақтауға қойылады. Темір хелатын күңгірт шыны ыдыста сақтау керек.
Витаминдердің концентрленген ерітінділерін әрқайсысын жеке шыны ыдыста сақтаймыз. Ерітінді дайындау үшін он еселенген витамин үлгісін алып, 10мл суда ерітеміз. 1мл витамин ерітіндісінде 1л Мурасиге-Скуга ортасына қажетті мөлшері болады.
Мурасиге-Скугтың сұйық және агар қосылған қоректік орталарын дайындау әдісі
Өсімдік жасушаларын in vitro жағдайында осіргенде қоректік орта кұрамын дұрыс таңдап алудың маңызы зор, себебі олар гетеротрофты коректенеді.
Қоректік ортаны таңдау өсімдіктің түріне және эксперименттің мақсатына байланысты. Әдетте Мурасиге мен Скугтың, Шенк -Хильдебрандтың, Гамборгтың В5, Уайттың, Хеллeрдің, Линсмайер -Скугтың қоректік орталарын пайдаланады. Бұл орталар даражарнакты және косжарнақты өсімдіктердің әр алуан түрлерін in vitro жағдайында өсіруге жарамды болып табылды.
in vitro жағдайында өсімдік жасушалары мен ұлпаларын өсіруде сұйық және қатты орталарды қолданады.Сұйық орталар суспензияларды, каллусты, оқшауланған денелер мен ұлпаларды регенерант өсімдіктерін культивирлеу үшін пайдаланады. Бұл үшін пробиркалаларға синтетикалық сілтілеу материалдарды немесе сүзгіш қағаздардан арнайы ұстап тұрғыш орналасады.
Қатты (агарланған) ортаны 5,6-6-рН-ында сумен гелді құрайтын теңіз балдырлары құрамына кіретін агар-агармен-полисахарид негізінде дайындайды, ал кейде тығыздығын және орын алмастырғыш ретінде Рю, Ріоо (биогель) полиакриамильді гельдерін қолданады. Агар-агар құрамында басқа заттар да бар, сондықтан оны 2-4 сағат бойы суды жиі өзгертіп жуу керек. Өсіруге алынатын ұлпалардың қажетіне қарай қоректік ортадан агар-агардың мөлшері 0,8 ден 1,5 % -ке дейін болады.
Жасушаларды сұйық қоректік ортада өсіру немесе суспензияда өсіру деп, қалқыған жасушаларды немесе кішігірім жасушалар тобын аппаратура арқылы таза ауамен қанықтырып және араластыра отырып сұйық қоректік ортада өсіруді атайды. Сұйық ортада жасушалардың таза ауамен қанықтырылуының әр түрлі тәсілдері бар: ыдысты тоқтаусыз айналдыру немесе шайқау, тағы бір жолы сұйық ортаға залалсыздандырылған ауа үрлеу арқылы. Жасушаларды сұйык қоректік ортада өсірудің өзіндік артықшылықтары бар: олар коректік заттармен жақсы қамтамасыз етіледі және токсинді заттары тез бөлініп шығады. Ұлпаларды қатты немесе сұйық орталарда болсын өсіргенде, ортаның рН-ы маңызды рөл атқарады, ұлпаларды өсіруде жетістікке жету үшін орта рН-ы қолайлы және тұрақты болу керек.
Макромолекулалардың активтілігі мен кұрьлымы ортаның рН-на тәуелді, әсіресе ұлпаның құрамындағы ферменттік белоктарға әсері жоғары. Сонымен бірге, ортаның рН-ы қоректік ортаның компоненттерінің тұрақтылығымен оларды жасушалардың қабылдап алуына, әсіресе өсуді реттегіштер мен витаминдердің сіңуіне әсер етеді. Егер ортаның рН-ы төмен болса, агар қатпайды, қоректік ортаның рН-ы белгілі деңгейге дейін, қышқыл немесе сілті косып жеткізіледі. Көптеген ұлпаларды өсіргенге орта рН-ы 5,5 -5,8 тең болу керек.
Қоректік орталарды дайындағанда және жасушаларды өсіргенде кейбір физикалық факторларды ескеру керек, мысалы: қоректік ортаның осмостық қысымы, температура және жарық. Жоғары осмостық қысым қоректік заттардың сіңуін қиындатады. Температура метаболизм процесстеріне әсер етеді, көптеген өсімдіктерді өсіру үшін ең қолайлы температура 24-27 градус.
2.3. Өсімдік материалын дайындап, эксплаптты бөліп алу әдісі
Эксплант өсімдіктердің әртүрлі мүшелерінен алынады, мысалы тұқымынан, тұқым жарнағынан, жапырағынан, сабағынан, ескінінен, бүршігінен және т.б. Эксплант -дегеніміз in vitro жағдайында өсіруге алынған өсімдіктің бөлігі. Эксплант алу үшін міндетті түрде таза, сау өсімдікті таңдап алу кажет.
Өсімдік материалын дайындап, эксплантты бөліп алу үшін алдын-ала дайындау жүргізудің маңызы зор. Өсімдік материалдарын дайындаудағы негізгі іс-шара оларды залалсыздандыру болып табылады. Өсімдік ұлпалары мен жасушаларын есіруде, микроклональді көбейтуде, каллусты және ісікті жасушалар алуда, гормональді реттелуді меңгеруде залалсыздандырылған өсімдік экспланттары пайдаланады.
Тұқымды өсіру үшін суға немесе қоректік ортаға отырғызады. Өсімдік түрлерін залалсыздандыру алдын жаксылап ағынды сумен, кейде жуғыш затгармен жуып алады, артық заттардан тазартады. Тамыртүйнектер мен тамырлардың қабығын аршиды, потоген -қабыршағын, бұтактарынан жабын қабыршақтарын тазартады.
Өсімдік экспланттарын сулеманың сулы ерітіндісінде немесе екіхлорлы сынып ерітіндісінде (0,1%), бромда (1%), пергидрольда (20-30%), хлораминде (3-6%), диацидте, натрий гипохлоридтінде (10%) антибиотиктерді құрайтын ерітінділермен залалсыздандырады.
Сулема уытты зат, оны сақтау кезінде және жеке түрлер үшін қажетті концентрацияны таңдағанда да ерекше сақтықты қажет етеді. Ұрыктарды 0,1 процент ерітіндіде 1-3 минут бойында, ал тамырды және түйнекжемістерді 10-20 минутқа дейін залалсыздандырады.
Активті хлор кұраушы ерітінділерді 1 рет қолданады және оларды жұмыс алдында ғана дайындайды.
Тамыржемістерін, ұрықтарды, ұлпа түйірін, оқшауланған ұрыктарды, тозаңдарды залалсыздандыру үшін диацидтің 0,2 процент ерітіндісі пайдаға асырылады. Диацидті этаномеркурхлоридінің 330 мг және 660 мг хлорид-цеталпиридинді ыстық суда (330 мл) бөлек еріте отырып дайындайды, содан соң оларды аралыстырып сұйық көлемін 1л-ге дейін жеткізеді де гвин детергенты - 80-нің бірнеше тамшысын қосады; оны қараңғыда қатты жабылған ыдыста (колбада) сақтайды.
Антибиотиктерді инфицирленген бактериялармен өсімдік материалын залалсыздандыру үшін пайдаланады. Көбінесе стептомицинді және 10-80 мг/л - тетрамицинді, 200-400 мг/л ампицилинді, левомицинді және т.б. қолданады.
Дайын залалсыздандырылған өсімдік экспланттарын алудың бірнеше тәсілдері бар, тәсілдер мүшелер түрлеріне тікелей тәуелді.
Жапырақтан эксплантты бөліп алу. Алдымен жапырақтан жүйкесін аламыз да жоғарыда көрсетілген тәртіппен залалсыздандырамыз. Стерильденген, залалсыздандырылған жапырақты ламинар-бокс ішінде стерилденген Петри табақшасына салып, стерильді пинцет және № 11 скальпель көмегімен 5-10мм көлемінің шамасында эксплантты кесіп алып, өсіру мақсатына байланысты қоректік ортаға ауыстырамыз (1- сурет).
Өскіннен эксплантты бөліп алу. Тұқымды алдын- ала таза стерильді жағдайда өсіріп, өскін аламыз. Стерильді өскінді ұзындығы 1 см ұзындықта ламинар-бокс ішінде стерильді Петри табақшасында тұқымнан кесіп аламыз. Өскінді жеке стерильді Петри табақшасына салып, стерильді пинцет және № 11скальпель көмегімен 2-3 мм көлемінің шамасында эксплантты кесіп алып, өсіру мақсатына байланысты қоректік ортаға ауыстырамыз.
Тұқымдардан эксплантты бөліп алу. Тұқымды тиісті әдістеме бойынша стерильдейміз де ісіндіруге (тұқымның ерекшелігіне байланысты 12 сағаттан 3 тәулікке дейін) ламинар-бокске қоямыз. Ісінген тұқымның қабығынан ажыратамыз. Қайтадан натрий гипохлоритте 20минут бойы стерилдейміз. Дистилденген сумен үш рет шаямыз да жаңа Петри табақшасына қойып, стерильді пинцетпен ұстай отырып стерильді скальпель көмегімен тұқымжарнақ көрінетіндей етіп кесеміз. Тұқымжарнақтың ұшынан 2-3 мм эксплант алып, өсіру мақсатына байланысты қоректік ортаға ауыстырамыз.
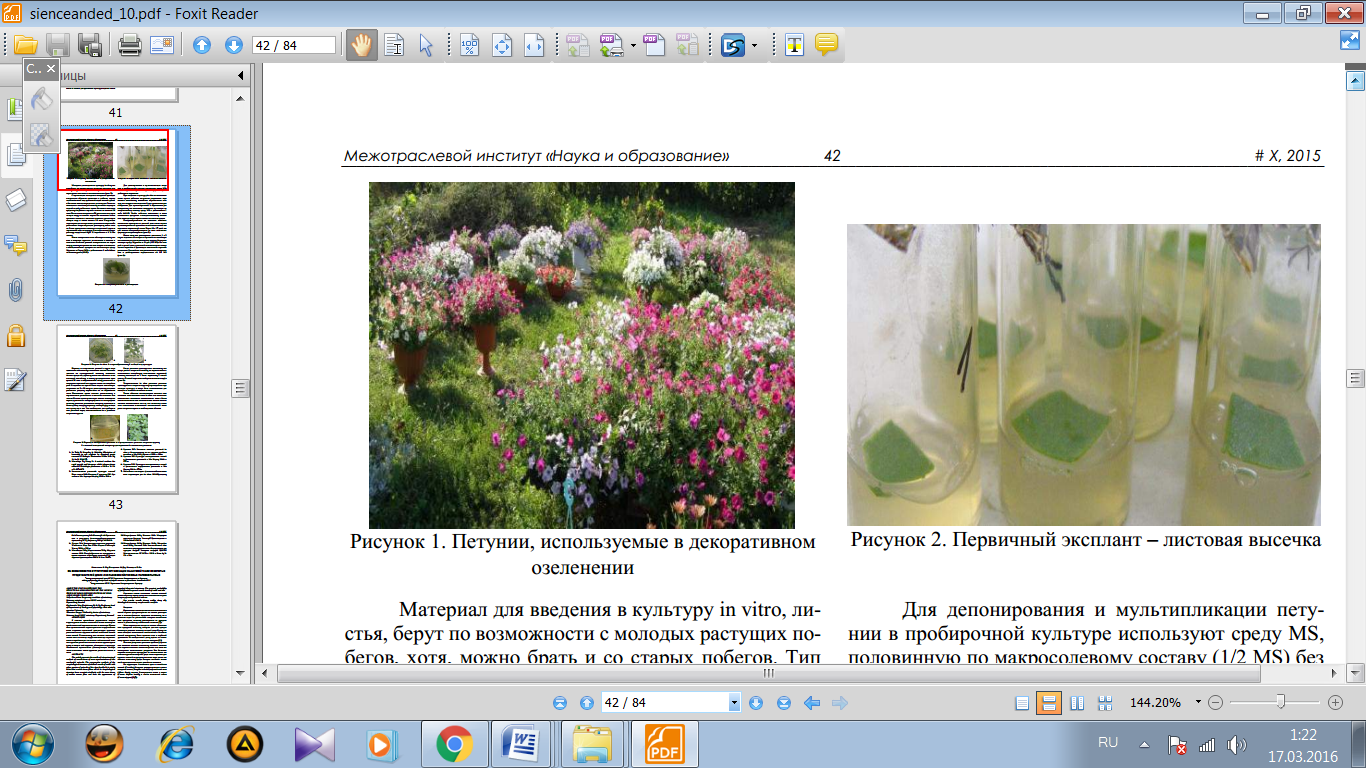
1- сурет. Жапырақты in vitro өсіру.
2.4. Эксплантты агары бар ортаға отырғызу және өсіру әдісі
Өсімдік мүшесінің кесіндісі, яғни эксплант (эксплантант), лайықты қоректік ортаға отырғызылса, біраз уақыт өткен соң оның кұрамындағы жасушалар бөлініп, өсе бастайды. Нәтижесінде каллус ұлпалары пайда болады.
Өзіне тән бір қалыпты өсуімен және көбеюімен сипатталынатын ретсіз өсетін каллус жасушалары қайта дифференциялану процестері арқасында, ұлпалар (гистогенез), мүшелер (органогенез) және ұрық тәріздес құрылымдар-эмбриондар (эмбриогенез, немесе сомалық эмбриогенез) түзе алады. Қоректік ортаның құрамын өзгерте отырып, ретсіз өсіп жатқан ұлпалардан өсімдіктің тотипотенттілік қасиеті арқасында регенерант- өсімдік алуға болады. Каллус жасушалары өсу қарқындьлығымен ерекшеленеді, түсі арқылы (ақ- ақшылдан, қара -қоңырға дейін, тығыздығына қарай) бос-борпылдақ, тығыз, т.б. түрлері, жарықта түсін өзгертіп көгеретіндігімен тағы сол секілді белгілерімен бір-бірінен ажыратылады.
Бастапқы кезде каллус өсіп, массасы қандай да бір ауыспалы кезеңге жеткеннен кейін морфогенезге ауысады. Бірақ, тек бірлі жарым санаулы жасушалар ғана, өзінің даму бағдарламасын өзгертеді.
Қоректік ортада міндетті түрде фитогормондар (соның ішінде ауксиндер) болған жағдайда жасушалар дедифференцияланып каллустың жасушаларға айналады. Маманданған жасушаның дедифференциялануының бірінші кезеңі ол жасушаның бөліну қабілетінің қалпына келуі.
Маманданған жасушаның өз қасиетін жоя бастағаннан (дедифференциялану) кейін 6-12 сағ. өткен соң олар жасушаның нәзік кұрылысында болатын өзгерістер болады. Жасуша қабығы жұмсарып, ісінеді, рибосомалар саны, эндоплазмалық тор және Гольджи аппараты элементтерінің саны өседі, ядрошықтың көлемі және саны көбейеді. Жасушаларда тРНҚ және рРНҚ синтезі артып, ДНҚ синтезі басталады, жаңадан түзілген белоктардың мөлшері көбейеді. Осы өзгерістердің барлығы, одан кейінгі жасушаның белінулері тек ортада ауксиндер мен цитокининдердің қатысуымен ғана жүреді. Жасуша бөлінулері 48-72 сағат өткеннен соң басталады.
Жасушалардың көбею стимуляторы /ауксин/ әсерінен сыртқы мембрана плазмалемманың өткізгіштігі артады да, оның қасиеттері өзгереді. Осыдан соң бұрыннан бар болған немесе жаңадан синтезделген матрицалық РНҚ-да цитоплазмалык ақуыздардың түзілу барысы активтенеді. Түзілген ақуыздар цитоплазмадан ядроға өтеді, арнайы гистондық белоктармен ерекше байланыска түседі де, транспорттық және рибосомдық РНҚ-лары синтезделуіне жауапты локустарды іске қосады. Жасушадағы РНҚ-ның мөлшері белгілі бір сатылы деңгейге жеткенде ДНҚ-ның репликациясы басталады. Ортада кинетин болмаған жағдайда жасушаның митозға өтуі болмайды. Кинетин жасушаның митозға өтіп бөлінуін аяқтау үшін қажет, арнайы РНҚ-лары мен белоктардың синтезделуін белгіледі. Жасушаның бөлінуіне қажет барлық процесстер жүру үшін, ауксиннен кейін кинетиннің әсері болу керек немесе екеуінің қосылған әсері қажет.
Жасушалардың өсуі ішкі және сыртқы ықпалдарға (фактор) тікелей байланысты болып келеді. Ішкі ықпалдарға жататындар: пролиферациялық қоры; жасушаның созылу ұзақтығы; циклға енген жасушаның жалпы күйі; жеке жасушалардың бөліну саны. Пролиферациялық қоры деп бөлініп жатқан жасушалар санының жалпы жасушалар санына қатынасын атайды. Бұл ара қатынасты пайызбен белгілесе, онда ол митоздық индекс деп аталады.
Сыртқы факторларға жататындар: қоректік ортаның құрамы, рН-тың деңгейі, оттегінің мөлшері, температура, отырғызылған жасушалар мен ұлпалардың тығыздығы, т.б.
Каллус ұлпалары өсімдіктің әртүрлі мүшелерінен әртүрлі жасында алынады.
2.5. Асептикалық өскіндерден каллус ұлпаларын алу әдісі
Құрамында фитогормондары бар қолайлы қоректік ортаға отырғызылған өсімдік ұлпалары дедифференцацияланып, бөліне бастайды да каллус ұлпаларын түзеді. Каллус ұлпаларын өсімдіктің әртүрлі мүшелерінен басқа асептикалық өскіндерден де алуға болады. Каллусогенез процесі бастапқы экспланттың көлеміне, жасына, оның өсімдікте орналасқан жеріне және пайдаланылған коректік ортаға байланысты болады. Өсімдіктердін тұқымжарнағын, жапырағын және гипокотильін қолайлы коректік ортада өсіргенде төмендегідей морфогенетикалық өзгерістерге ұшырайды.
1.Эксплант бастапқы каллусты түзіп, одан әрі адвентивтік бүршік пайда болады;
2.Түзілген каллустың бастапқы және ауыстырмалы ұлпаларынан меристемалык бастамалар түзіледі;
in vitro жағдайында түзілген қиярдың каллус ұлпалары алынған бастапқы эксплантына байланысты, әдетте қара-жасыл, қою-сары және сары-жасыл түсті болып келеді. Консистенциясына байланысты созылмалы, борпылдақ және таңба тәрізді болады.
Өсімдік жапырағынан каллус ұлпаларын алу
Жапырақ жасушаларын in vitro жағдайында қолайлы қоректік ортаға отырғызылғанда бұрынғы атқаратын қызметі мен құрылысын, құрылымын өзгертіп, дифференцацияланып бөліне бастайды, өте тез өсетін каллус ұлпаларын түзеді. Жасушаларды қараңғы жағдайда өсіргенде жапырақ өзінің жасыл түсін өзгертіп, фотосинтездік қабілеттілігін жоғалтады. Жапырақтан каллус алу үшін ортаңғы жүйкелердің аймағы өте қолайлы болып табылады. Каллус алу үшін стерильді пробиркалық өсімдік және стерильді емес жапырақтар пайдаланады (2- сурет).
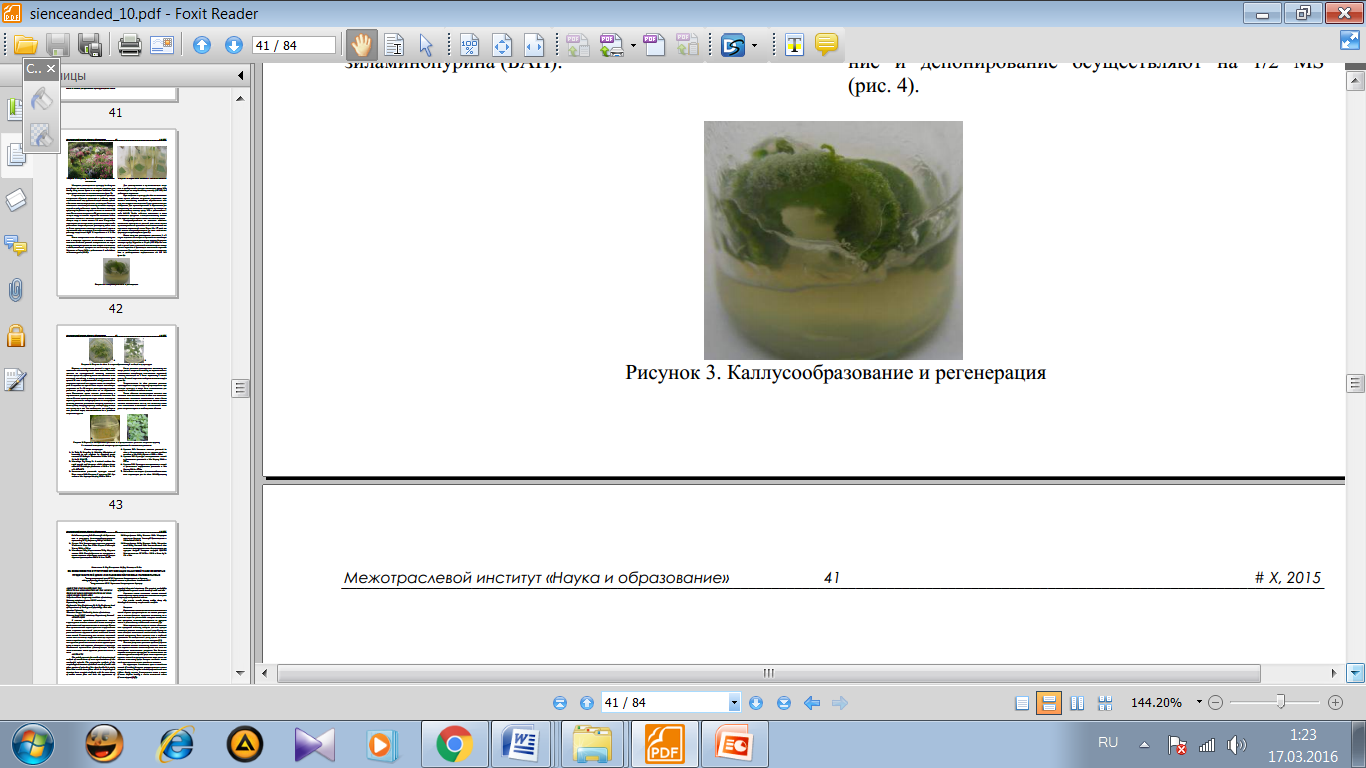
2- сурет. Өсімдік жапырағының in vitro алынған каллусы
2.6. Өсімдіктерді микроклонды көбейту және отырғызу әдісі
Маманданған ұлпаның кез келген тірі жасушалары лайықты қоректік ортада өсіргенде, өздерінің тотипотенттік қасиетін жүзеге асырып, регенерация арқылы бүтін өсімдікке айнала алады. Жеке жасушалардан сол өсімдік түріне тән барлық белгілері мен қасиеттері сақталған бүтін өсімдіктің түзілуі клондық көбейту технологиясының негізін қалайды. Клон деген -жыныссыз жолмен, яғни вегетативтік көбею жолымен, түзілетін ағза.
Өсімдіктердің клондык микрокөбеюі деген өсімдіктердің in vitro жағдайында жыныссыз жолмен көбеюі. Соның нәтижесінде пайда болған өсімдіктер бастапқы өсімдікпен және өзара бір-бірімен генетикалык тұрғыдан айнымастай бірдей болады.
Бұл биотехнологиялық әдістің дағдылы вегетативтік жолмен көбеюмен салыстырғанда бірталай артықшылықтары бар, атап айтқанда: көбею коэффициенті өте жоғары; микрокөбеюмен қатар өсімдіктер вирустар мен патогендік микроорганизмдерден сауықтырылады; сұрыптау процесін жылдамдату, жаңа сорттарды тез көбейтіп, оларды ауылшаруашылык өндірісінде пайдалану мерзімі едәуір қысқарады; вегетативтік жолмен көбейе алмайтын өсімдіктерді in vitro жағдайында көбейтуге болады; үнемділік; жас өсімдіктерді алу, кәрі дарақтарды жасарту; өсу процесін жыл бойы үзбеуге болады, әсіресе бұл дамуында тыныштық кезеңі болатын өсімдіктерді көбейтуге тиімді.
Клондық микрокөбейту әдістері in vitro жағдайында қолтық бүршік меристемаларын өсіруге және басқа экспланттардан немесе каллустардан бүршіктер мен эмбриоидтарды өсіруге негізделген.
Клондық микрокөбейту жұмысы 4 кезеңнен өтеді.
І-кезең. Эксплантты in vitro жағдайында өсіру. Бұл кезде толығымен залалсызданған эксплантты алып, лайықты қоректік ортаға отырғызып, оның жақсы өсуіне қолайлы жағдай туғызу керек. Бұл кезеңнің нәтижелілігі эксплантты дұрыс таңдап алуға байланысты. Ол үшін донор өсімдіктің әр даму кезеңінде ұлпаларынан немесе мүшелерінен экспланттар алынып, олардың in vitro жағдайында өсу және морфогенездік қабілеттері зерттеледі.
II -кезең. Микрокөбейту. Өркендердің пайда болуы және олардың санын көбейту.
III-кезең. Өркендерді тамырландыру және оларды сақтау. Жақсы тамыр жүйесі пайда болуына және дамуына жағдай туғызу. Ол үшін қоректік ортаға ризогендік факторы, яғни ауксин қосылады. Тамырлар өсіп жетілген соң өсімдіктерді топыраққа көшіруге дайындайды. Төмен температура өсімдіктердің дамуын тежеп, ұзақ уақыт сақтауға мүмкіншілік береді.
ІV-кезең. Өсімдіктерді топыраққа отырғызу. Өсімдіктерді топыраққа отырғызу алдында оларды арнайы дайындайды. Сол үшін ауаның ылғалдылығын және жарықтың қарқындылығын арттырады. Өсімдіктер гетеротрофтық қоректенуден антотрофтык қоректенуге өтеді. Бұл ұқыптылықты талап ететін ең маңызды кезең. Клондық микрокөбейту нәтижесі өсімдіктің генотипіне, жасына, экспланттың тегіне, қоректік ортаға, өсіру жағдайларына байланысты.
Микроклональді көбейтуде тамыр түзуін индукциялау
Құрамында цитокинині бар қоректік ортада бүлдіргеннің апикальді меристемасын өсіргенде қолтык меристемаларының белсенділігі нәтижесінде бір төбе бүршіктер бірнеше сабақ бүршіктері пайда болады, өсу нүктелері ұлғайып, жаңа жапырақтары пайда болады. Бірақ бұл қоректік ортада тамыр пайда болмайды, ол үшін бүршіктерді құрамында ауксині бар жаңа қоректік ортаға отырғызу қажет.
Өсімдік материалдарын сауықтыру
Биотехнологиялық негізде вируссыз тұқым шаруашылығы - тауарлы шаруашылықтағы оның өнімділігі мен сапасын арттырудың ең тиімді жолы. Вируссыз тұқым шаруашылығы бірқатар сатылардан тұрады:
1. Сорттарды сауықтыру. Осы мақсатпен апикальді меристема әдісі немесе оның термо немесе химиотерапиямен байланысы кең көлемде қолданылады;
2. Зертханалық және жылыжайларда сауықтырылған өсімдіктерді жедел көбейту;
3. Біріншілік және таңдаулы тұқым шаруашылығымен
айналысатын
маманданған шаруашылықтардың қажеттілігін қанағаттандыратын
көлемде
далалық жағдайдағы меристемалық клондарды бір-екі жыл көбейту,
біріккен
клонды материалдардың зақымдалуын бақылау;
4. Жоспарланған көлемде супер-супер таңдаулы
және таңдаулы түрлерін
алғанға дейін өнімнің сапасын сақтауға және қайталанатын
инфекциядан
сақтауға бағытталған арнайы технологияны сақтау арқылы арнайы
тұқым
шаруашылығында сауықтырылған негізгі материалды көбейту;
Ұштық меристеманы оқшаулау
Меристеманы жасыл немесе этиолирленген өскіндерінен бөліп алады. Меристеманың жасыл өскіндерден түзілуі техникалық процесс, онда олар жақсы ажыратылады және түзілуі кезінде зақымдануға төзімдірек болады. Бірақ этиолирленген меристеманы қолданғанда сауықтыру жұмыстарының тиімділігі жоғары болады, себебі мұндай өскіндердегі вирустар концентрациясы айтарлықтай төмен, сондай-ақ вирусы жоқ ұштық меристема аумағы жарықта өсірілген өскіндердікінен әлде қайда көп болады. Меристема 2 см немесе одан үлкен болып өскеннен кейін оны бөліп алады.
Меристеманы өсіру жұмыстары сынапты-кварцті немесе БУВ типті бактериоцидті шамның көмегімен залалсыздандырылған микробиологиялық боксте жүргізіледі. Жұмысты бастар алдын жұмыс орнын (столды, бинокулярлы лупаны) және пробиркалары бар штативті спиртпен сүртеді. Өскіндермен жұмыс жасауға арналған құралдарды (пинцеттерді, скальпельдерді, инелерді) әрдайым қолданар алдында спиртке салып, содан соң от жалынына қару арқылы залалсыздандырып отырады.
ІІІ. КҮТІЛЕТІН НӘТИЖЕЛЕР
Ғылыми жұмыс мақсатына сай міндеттерді шешуде келесі нәтижелер күтіледі:
- Жоба шегінде өткізілген ғылыми зерттеу жұмыстарының нәтижелерін ҚР білім мен ғылым саласын бақылау комитеті тізіміне кіретін журналдарда, Халықаралық, шетелдік конференцияларда және Scopus немесе нольдік емес импакт факторы бар журналдарда басып шығару жоспарланған;
- Экологиялық факторлардың in vitro әдісімен өсірілген Petunia Juss. туысы түрлеріне әсерін, Арқалық қаласының климаттық жағдайына төзімділігі зерттелінеді;
- Арқалық қаласында Petunia өсімдігін in vitro әдісімен көбейту жолдарының мүмкіндіктері анықталады;
- Арқалық қаласын көгалдандыру мақсатында in vitro әдісімен Petunia туысы түрлерін жылдам көбейту, сапалы көшеттер өсіруге мүмкіндік тудырады;
- Ғылыми жұмыс нәтижесінде Арқалық қаласын көгалдандыру мекемесімен бірлескен ынтымақтастық жұмыс жүргізу;
- Арқалық қаласында жобаны жүзеге асыру нәтижесінде қаланы сәндік гүлдермен безендіру диапазоны кеңейтіледі;
ҚОРЫТЫНДЫ
Бүгінде өсімдіктерді жылдамдатып көбейту кезінде биотехнология әдістері кеңінен қолданылады, оның ішінде in vitro жағдайында қалемшелеу арқылы өсімдік регенеранттарын жедел көбейтіп өсіру кең көлемде қолданылу.
Ғылыми - техникалық прогрестің қарқынды дамуы өсімдіктерді көбейтудің жолдары жеңілдетілген және тездетілген жаңа технологияларды ауыл шаруашылығында қолдануға мүмкіндік туғызды. Сондай тиімді әдістердің бірі ретінде өсімдіктерді микроклональды жолмен көбейту болып табылады. Микроклониальді көбейту -«жабық тамыр жүйесі» технологияны және биостимуляторларды пайдаланып өсімдіктерді вегетативтік жолмен көбейту болып табылады, яғни in vitro жағдайында генетикалық біртекті дәл өзіндей көшірме алу. Петуния өсімдігін in vitro әдісімен көбейту дәстүрлі вегетативті көбейтуге қарағанда тиімді болы отыр. Іn vitro әдісімен көбейту арқылы аз уақыт аралығында қажетті мөлшерде генетикалық бір текті өсімдік материалын алуға мүмкіндік туды.
Қорыта келе, Ғылыми жұмыстың нәтижесінде Арқалық қаласында Petunia Juss.</<font face="Times New Roman, serif"> (Петуния) туысы түрлері in vitro әдісімен өсіріліп, қалада Petunia өсімдігінің сапалы көшеттерін көбейтуге мүмкіндік туады.
Пайдаланылған әдебиеттер тізімі
-
12. Валиханова Г. Ж., Рахимбаев И. Р. Культура клеток и биотехнология растений. Алма-Ата: Изд. КазГУ, 1989. 80 с.
-
Глеба Ю. Ю., Сытник К .М. Слияние протопластов и генетическое конструирование высших растений. Киев: Наукова думка, 1982. 104 с.
-
Глеба Ю. Ю., Сытник К. М. Клеточная инженерия. Киев: Наукова думка, 1984. 160 с.
-
Калишш Ф. Л., Сарнацкая В. В., Полищук В. Е. Методы культуры тканей в физиологии и биохимии растений. Киев: Наукова думка, 1980. 488 с.
-
Катаева Н. В., Бутенко Р. Г. Клоналное микроразмножение растений. М.: Наука, 1983. 97 с.
-
Культура клеток растений. М.: Наука, 1981. 167 с.
-
Кулыура клеток и биотехнология. М.: Наука, 1986. 285 с.
-
Муромцев Г.С., Бутенко Р. Г., Тихоненок Т. И., Прокофьев М, М. Основы сельскохозяйственной биотехнологии. М.: ВО Агропромиздат, 1990. 384 с.
-
Пирузян Э. С. Основы генетической инженерии растений. М.: Наука, 1988. 303 с.
-
Рахимбаев И. Р., Колумбаева С. Ж., Джокебаева С. А. Культура клеток и клеточная инженерия растений. Алматы: Изд. КазГУ, 1993, 80 сг
-
Сассон А. Биотехнологая: свершения и надежды. М.: Мир, 1987.411 с.
-
Уотсон Дж, Туз Дж., Курц Д. Рекомбинантная ДНК. М.: Мир, 1986. 285 с. .
-
Албертс Б., Брей Д., Люис Дж., Рэфф М., Робертс К. Уотсон Дж. Молекулярңая биология клетки Т. 2. М.: Мир 1986, С. 259- 281.
-
Аяаньев Е. В., Баширов В. Н. Генетическая инженерия растений: проблемы и перспективы//Биология наших дней. М.: Знание, 1987. С. 25-41.
-
Анапиев Б. Б. Морфогенез в культуре изолированных пыльников и микроспор пшеницы. Автореф. дис. канд. биол. наук. Алма - Ата, 1992. 25 с.
-
Батыгина Т. Б., БутенкоР. Г. Морфогенетические потенции зародыша покрытосеменных растений//Бот. ж., 1981, т. 66, №11. С. 1531- 1548.
-
Батыгина Б. Т., Васильева В. Е., Маметьева Т. Б. Проблемы
морфогенеза іn vitro. Эмбриогенез у покрытосеменных растений//Бот. ж., 1978, т. 63, №1. С. 87-95. -
Бишимбаева Н. К. Морфологическая гетерогенность и регенерационная способность каллусных клегок ячменя. Автореф. дис. канд. биол. наук. Душанбе, 1989. 22 с.
-
Бугара А. М., Русина Л. В. Культура неоплодотворенных завязей и семяпочек іп vitro как способ получения галловдных растений//Физ. и биох. культ. раст., 1988, т. 20, № 5. С. 419 -430.
-
Бутенко Р.Г. Выращивание клеток высших растений всуспензионной культуре // Известия АН СССР, сер. биол., 5,1977.С. 697-709.
-
Валиханова Г. Ж., Бабаев Р. А., Бишимбаева Я. К., Заирова А. С. Методическое руководство по применению ЭВМ в НИРС по культуре тканей. Алма-Ата: КазГУ, 1989. 31 с.
-
Гейзен Г., Диз П., Ваи Монтапо М., Шелл Й. Движение ДНК через генетические барьеры: влияние корончатых галлов// Мобилъность генома растений. М.: ВО Агропромиздат, 1990, С. 18 - 55.
-
Давоян Э. И. Получение гаплоидов у риса методом культивирования неоплодотворешшх завязей в условиях іп vitro //Доклады ВАСХНИЛ, №3, 1985. 45 с.
-
Джардемалиев Ж. К. Биотехнологические аспекты регенерации растений в культуре клеток и пршопластов пшеняцы. Автореф. дис. канд. биол. наук. Ташкент, 1994. 27 с.
-
Джори Б. М., Рао П. С. Экспериментальная эмбриология //Эмбриология растений: использование в генетике, селекции, биотeхнологии. Т.2.М.: ВО Агропромиздат, 1990. С. 343 - 427.
-
Дьячук Т. И., Дьячук П. А. Культура пыльников злаков: современное состояние, проблемы, перспективы. С.~ х. Биол.,1989, № 5. С. 3 - 10.
-
Зенгбуш П. Молекулярная и клеточная биология. Т.З. М.: Мир, 1982. 344 с.
-
Искакова К.М. Морфогенез в длительно поддерживаемой культуре андрогенных каллусов ячменя. Автореф. дис. канд. биол. наук. Алма - Ата, 1991. 21 с.
-
Карабаев М. Кулътивируемые клетки пшеницы и кукурузы: физиологические и биотехнологические аспекты. Автореф. дис. доктора биол. наук. М., 1994, 49 с.
-
Кударов Б. Р, Тивари Ш., Рахимбаев И. Р. Методические рекомендации по культуре пыльников и изолированных микроспор ячменя и пшеницы. М, 1990. 34 с.
42