- Учителю
- Явление изменчивости на примере видов Ульяновской области
Явление изменчивости на примере видов Ульяновской области
-
Изменчивость.
-
Модификационная изменчивость.
Модификационная изменчивость - это эволюционно закрепленные реакции организма на изменения условий внешней среды при неизменном генотипе. Такой тип изменчивости имеет две главные особенности. Во- первых, изменения затрагивают большинство или все особи в популяции и у всех них проявляются одинаково. Во-вторых, эти изменения обычно имеют приспособительный характер. Как правило, модификационные изменения не передаются следующему поколению. Классический пример модификационной изменчивости дает растение стрелолист, у которого надводные листья приобретают стреловидную форму, а подводные - лентовидную.
Если у гималайского кролика на спине удалить белую шерсть и поместить его в холод, на этом месте вырастет черная шерсть. Если черную шерсть удалить и наложить теплую повязку, вырастет белая шерсть. При выращивании гималайского кролика при температуре 30*С вся шерсть у него будет белая. У потомства двух таких белых кроликов, выращенного в нормальных условиях, появится "гималайская", окраска. Такая изменчивость признаков, вызванная действием внешней среды и не передающаяся по наследству, называется модификационной.
Обычно, говоря о модификационных изменениях, имеют в виду морфологические изменения (например, изменение формы листьев) или изменения окраски (некоторые примеры приведены в п. Влияние генотипа и среды на фенотип ). Однако нередко в эту группу включают и физиологические реакции. Регуляция работы генов лактозного оперона кишечной палочки представляет собой пример такой физиологической реакции. Напомним, в чем она состоит. При отсутствии в среде обитания бактерий глюкозы и при наличии лактозы бактерия начинает синтезировать ферменты для переработки этого сахара. Если же в среде появляется глюкоза, эти ферменты исчезают и бактерия возвращается к стандартному обмену веществ.
Другой пример физиологической реакции - увеличение числа эритроцитов в крови у человека, поднявшегося в горы. Когда человек спускается вниз, где содержание кислорода нормально, число эритроцитов возвращается к норме.
В обоих примерах модификационные изменения имеют ясно выраженный приспособительный характер, поэтому их часто называют физиологическими адаптациями.
Модификационная изменчивость не затрагивает наследственной основы организма - генотип и поэтому не передается от родителей потомству.
Еще одна особенность модификационной изменчивости - ее групповой характер. Определенный фактор внешней среды вызывает сходное изменение признаков у всех особей данного вида, породы или сорта: под воздействием ультрафиолетовых лучей все люди загорают, все растения белокочанной капусты в жарких странах не образуют кочана. При этом, в отличие от мутаций, модификации направленны, имеют приспособительное значение, происходят закономерно, их можно предсказать. Если листья на деревьях уже распустились, а ночью были заморозки, то утром листья у деревьев примут красноватый оттенок. Если мышей, которые жили на равнинах вблизи гор, переселить в горы, то у них повысится содержание гемоглобина в крови.
Благодаря возникновению модификаций особи непосредственно (адекватно) реагируют на изменение условий среды и лучше приспосабливаются к ней, что дает возможность выжить и оставить потомство.
Не все модификации обязательно адаптивны. При интенсивном действии многих агентов наблюдаются ненаследуемые изменения, случайные по отношению к вызвавшему их воздействию. Они проявляются только в условиях, которые их вызывают. Причины появления таких фенотипически измененных клеток связаны с ошибками процесса трансляции , вызванными этими агентами.
Таким образом, модификационная изменчивость не затрагивает генетической конституции организма, т.е. не является наследственной. В то же время она вносит определенный вклад в процесс эволюции. Адаптивные модификации расширяют возможности организма к выживанию и размножению в более широком диапазоне условий внешней среды. Возникающие в этих условиях наследственные изменения подхватываются естественным отбором и таким путем происходит более активное освоение новых экологических ниш и достигается более эффективная приспособляемость к ним.
Ход работы:
А) сбор листов Земляники ( 100 экземпляров);

Б) измерение длины и подсчет числа зубчиков средней доли листа земляники;
В) Статистическая обработка полученных результатов.
Результаты:
А) сбор листов Земляники:
Б )
измерение длины и подсчет зубчиков:
)
измерение длины и подсчет зубчиков:
 Максимальное
значение
Максимальное
значение
 Минимальное
значение
Минимальное
значение
Среднее значение
Вариационный ряд по 1 признаку ( количество зубчиков): F -
частота встречаемости.
Х - значение вариаций.
Графическое изображение вариационного ряда:
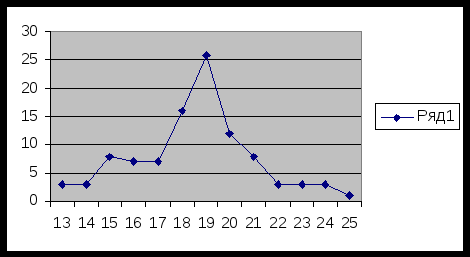
Исходя из результатов можно сделать вывод, что не все вариации встречаются одинаково часто, максимальная частота составляет 26.
Расчеты:
1) Х= А + Σ аf/n
X= 19 + (-46)/ 100= 18, 54 зуб.
Вариация очень близкая к X на графике находится очень близко к среднему значению. Стоит посередине ряда и встречается с максимальной частотой.
2) σ = Σа2f - (Σaf)2/100/ n
Стандартное отклонение. Показывает, на сколько в среднем отличается каждая из вариаций от Х.
σ = 648 - (-46)2/100 /100= 2,503
Стандартное отклонение составляет 2,503.
3) V= σ/x * 100%= 2, 503/ 18,54 *100%= 13,5%
Доля σ от Х.
4) t = X min - X/ σ= 13 - 18,54/ 2,503= 2,21
t = X max - X/ σ= 25 - 18,54/ 2,503= 2,58
Значение t показывает на сколько сигм отличаются минимальная и максимальная вариации от среднего арифметического в вариационном ряду. Все вариации укладываются в пределы 3 σ . Сюда попадает 99,74 % особей.
5) m (x)= σ/ n= 2, 503/ 100= 0,2503 ( зуб.)
Ошибка от среднего составляет 0,2503 зуб. На столько мы ошиблись, когда считали, что среднее арифметическое выборки соответствует среднему арифметическому генеральной совокупности.
-
Вариационный ряд по второму признаку ( по длине листа):
Lim = 6,6 -3,0/10= 0, 4 ( класс. Промежуток)
Графическое изображение вариационного ряда:
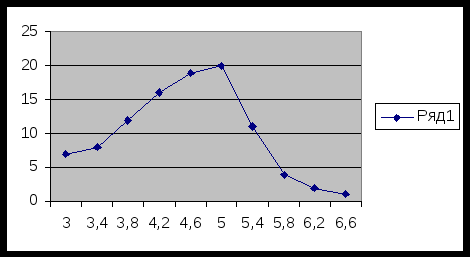
Вывод: максимальная частота встречаемости составляет 20 с вариацией 5,0-5,3.
Х составляет 5,0.
Расчеты:
1) X = A + λ * Σ a/f / n
X = 5,0 + 0,4 * ( -125) / 100 = 4,5 см
Среднее значение X на графике очень близко к среднему значению. Стоит посередине ряда и встречается с максимальной частотой.
2) σ = λ Σ ( a/ )2 - Σ (a/f )2/ n / n
σ = 0,4 * 555 - (-125)2/100 /100= 0,798 (см)
Стандартное отклонение. Показывает, на сколько в среднем отличается каждая из вариаций от Х.
t = x - x/ σ = 6, 6 - 4,5/ 0,798= 0,961
t = X - X/ σ = 3,0 - 4,5/ 0, 798= 2, 639
Значение t показывает на сколько сигм отличаются минимальная и максимальная вариации от среднего арифметического в вариационном ряду. Все вариации укладываются в пределы 3 σ . Сюда попадает 99,74 % особей.
v= σ / X * 100%=0,798/ 4,5 * 100%= 17,7% Доля σ от Х.
m (x)= σ / n=0, 798/ 100=0,0798 (см)
Ошибка от среднего. На столько мы ошиблись, когда считали, что среднее арифметическое выборки соответствует среднему арифметическому генеральной совокупности.
2.Мутационная изменчивость
Разнообразные формы и проявления модификационной изменчивости не
затрагивают генотипа организма. Наряду с модификациями существует
форма изменчивости, меняющая генотип. Эту форму изменчивости
называют генотипической или мутационной, а отдельные изменения -
мутациями.
Существование наследственных изменений было известно Дарвину. Вся
его теория эволюции вытекает из учения о естественном отборе
наследственных изменений. Наследственная изменчивость - необходимая
предпосылка естественного и искусственного отбора. Однако во
времена Дарвина еще отсутствовали опытные данные по
наследственности и законы наследования не были известны. Это не
давало возможности строго различать разные формы изменчивости в
зависимости от наследования.
Понятие мутаций было введено в науку голландским ботаником де
Фризом. Мутации затрагивают разнообразные стороны строения и
функции организма. Например, у дрозофилы известны мутационные
изменения формы крыльев (вплоть до полного их исчезновения),
окраски тела, развития щетинок на теле, формы глаз, их окраски
(красные, желтые, белые, вишнёвого цвета и т. п.), а также многих
физиологических признаков (продолжительность жизни, плодовитость,
стойкость к разным повреждающим воздействиям и т. п.).
Первоначальные представления де Фриза о том, что мутации всегда
крупные наследственные изменения, дальнейшими исследованиями не
подтвердились. Наряду с резкими отклонениями гораздо чаще
встречаются небольшие мутации, лишь немногим отличающиеся от
исходных форм. Тем не менее, указанный еще де Фризом признак
мутаций - их скачкообразный характер и наследственность - остается
в силе. Мутации совершаются в различных направлениях, и обычно они
не являются приспособительными, полезными для организма
изменениями.
Существуют и такие наследственные изменения, которые в гомозиготном
состоянии вызывают гибель (такие мутации называются
летальными).
Генные мутации. Различают несколько типов мутаций по характеру
изменений хромосомного аппарата. Наиболее распространенными
являются мутации, не связанные с видимыми в микроскоп изменениями
строения хромосом. Такие мутации представляют собой качественные
изменения отдельных генов и носят название генных мутаций. На
основании исследований, проведенных главным образом на
микроорганизмах за последнее время, установлено, что такие мутации
связаны с преобразованием химической структуры ДНК, входящей в
состав хромосом. Последовательность оснований определяет состав
образующейся на ДНК молекулы РНК, а она в свою очередь
обусловливает последовательность аминокислот при синтезе белковой
молекулы. Химическая основа генных мутаций заключается в изменении
расположения нуклеотидов в цепочке ДНК.
Хромосомные мутации. Наряду с генными известен ряд других мутаций,
связанных с видимыми преобразованиями хромосом, которые доступны
непосредственному микроскопическому изучению. К числу таких
изменений относится, например, переход части одной хромосомы на
другую, ей не гомологичную, поворот участка внутри хромосомы на
180° и ряд других структурных изменений отдельных хромосом.
Особую группу мутаций представляют собой изменения числа хромосом.
Эти мутации сводятся к появлению лишних или утере некоторых
хромосом. Такого рода изменения в хромосомном составе происходят
при нарушении в силу каких-либо причин нормального хода мейоза,
когда вместо нормального распределения хромосом между полюсами
ахроматинового веретена и затем дочерними клетками обе гомологичные
хромосомы отходят к одному полюсу. Обычно такого рода нарушения
оказываются неблагоприятными, снижающими жизнеспособность.
Особый тип наследственных изменений представляет собой явления
полиплоидии, которое выражается в кратном увеличении числа
хромосом. Возникновение полиплоидов обычно связано с нарушением
процессов митоза или мейоза. Эти нарушения сводятся к тому, что
хромосомы проделывают нормальный митотическйй цикл, а веретено
деления, служащее для «растаскивания» хромосом к противоположным
полюсам, не функционирует. В результате хромосомы не расходятся к
полюсам и не образуют дочерних ядер, а остаются в том же ядре. Если
этот процесс имеет место в соматической клетке с диплоидным набором
хромосом, то сразу возникает клетка с удвоенным диплоидным набором
(такие клетки называются тетраплридными). Они имеют, следовательно,
вместо двух гаплоидных наборов четыре (4 n). Если это наблюдается
при мейозе, то конъюгирующие гомологичные хромосомы не расходятся к
противоположным полюсам и возникают диплоидные гаметы. Если такая
гамета при оплодотворении сольется с нормальной гаплоидной, то
возникает триплоидная зигота (с тройным набором хромосом- 3 n).
Если же обе гаметы окажутся диплоидными, то возникает тетраплоидная
зигота.
Полиплоидные виды довольно часто наблюдаются у растений и очень
редко встречаются у животных. Полиплоидные мутации у растений по
сравнению с диплоидами часто характеризуются более мощным ростом,
большим размером и весом семян и плодов и т.п. В
генетико-селекционной работе по созданию высокопродуктивных сортов
растений явление полиплоидии широко используется на практике.
В настоящее время разработаны методы, позволяющие получать
полиплоиды экспериментально, воздействуя на делящуюся клетку
некоторыми ядами (например, колхицином), разрушающими веретено, но
не препятствующими удвоению хромосом.
Мы видим, таким образом, что мутации всегда связаны с изменениями в
хромосомах. Если эти изменения происходят в половых клетках, то они
проявляются в том поколении, которое развивается из половых клеток.
Но изменения могут иметь место и в соматических клетках. Тогда они
получают название соматических мутаций. Такие мутации приводят к
изменению признака только части организма, развивающегося из
измененных клеток.
Результаты:
Изменение окраски цветка

Изменение окраски цветка у Цикория обыкновенного


Мутация - пестролистность у Фиалок.


Махровость цветков у Фиалки.

Четырехлистность Клевера ползучего.


-
Закон гомологических рядов в наследственной изменчивости.
Гомологических рядов закон изменчивости, разработанный советским учёным Н. И. Вавиловым закон, устанавливающий параллелизм в изменчивости организмов. Ещё Ч. Дарвин (1859-68) обратил внимание на далеко идущий параллелизм в изменчивости близких видов и родов животных и растений. В 19 и начале 20 вв. ряд ботаников и зоологов специально изучали параллельную изменчивость разных видов растений и животных. Советский генетик Ю. А. Филипченко подытожил (1922) ряд таких, преимущественно зоологических, данных в статье о параллелизме изменчивости в живой природе, который он объяснял систематической и филогенетической близостью родов и видов, входивших в изучаемые группы.
Лишь Н. И. Вавилов подошёл к проблеме параллелизма в изменчивости близких видов и родов с генетических позиций и на основе сравнительного изучения обширнейшего мирового материала (в природных условиях, культурах и в опытах) по изменчивости ряда семейств растений, богатых хорошо изученными культурными видами, - главным образом злаков. Это позволило ему в 1920 на 3-м Всероссийском съезде селекционеров в Саратове выступить с докладом «Закон гомологических рядов в наследственной изменчивости». Н. И. Вавилов показал, что если все известные у наиболее изученного в данной группе вида вариации расположить в определённом порядке в таблицу, то можно обнаружить и у других видов почти все те же вариации изменчивости признаков. Более того, по мере развития исследований видов, входящих в данную группу, «пустые» места в таблице заполняются и параллелизм в изменчивости близких видов становится всё более полным. Принципиально сходный, но слабее выраженный параллелизм характеризует изменчивость различных родов в пределах семейства, и ещё менее полный - различных семейств в пределах группы более высокого ранга.
Таким образом, закон гомологических рядов сводится к следующему: близкие виды благодаря большому сходству их генотипов (почти идентичные наборы генов) обладают сходной потенциальной наследственной изменчивостью (сходные мутации одинаковых генов); по мере эволюционно-филогенетического удаления изучаемых групп (таксонов), в связи с появляющимися генотипическими различиями параллелизм наследственной изменчивости становится менее полным. Следовательно, в основе параллелизмов в наследственной изменчивости лежат мутации гомологичных генов и участков генотипов у представителей различных таксонов, то есть действительно гомологичная наследственная изменчивость. Однако и в пределах одного и того же вида внешне сходные признаки могут вызываться мутациями разных генов; такие фенотипические параллельные мутации различных генов могут, конечно, возникать и у разных, но достаточно близких видов. Н. И. Вавилов подчёркивал, что закон гомологических рядов неизбежно обнимает и такую, в генетическом смысле не строго гомологичную, фенотипически же параллельную изменчивость.
После 1920 представители школы Н. И. Вавилова в СССР, а также ботаники и селекционеры зарубежных стран накопили огромный фактический материал, подтверждающий всеобщность закона.
Сначала исследования касались в основном морфологических признаков; затем они были распространены на биологические, физиологические и биохимические свойства. Многочисленные подтверждения закона были получены на простейших, низших растениях, большом числе семейств высших растений и на животных.
Закон отражает всеобщее и фундаментальное явление в живой природе. Он имеет огромное практическое значение в растениеводстве и селекции, а также в животноводстве. На основе этого закона растениеводы и животноводы могут целенаправленно искать и находить нужные признаки и варианты у различных видов в почти бесконечном мировом многообразии форм как культурных растений и домашних животных, так и у их диких родичей. Эти поиски, особенно среди культурных растений и их диких предков, значительно облегчаются учением Н. И. Вавилова (1926 и др.) о центрах происхождения культурных растений и его работами (1927, 1928, 1930) о географических закономерностях в распределении генов культурных растений. Г. р. з. Н. И. Вавилова уже с 30-х гг. 20 в. явился мощным стимулятором целенаправленной селекции, создания новых сортов культурных растений и разработки научных основ интродукции и акклиматизации</<font size="4">. Закон играет всё большую роль в изучении механизмов эволюционного процесса, в истолковании ряда биогеографических явлений и в разработке основ современной систематики низших таксонов.
Семейство Бобовые:


Астрагал сладколистный Астрагал нутовый
Семейство Бобовые:


Горошек посевной Горошек лесной
Семейство Бобовые:


Люцерна посевная Донник лекарственный
Клевер ползучий
Семейство Розоцветные:


Яблоня домашняя Груша обыкновенная
Семейство Розоцветные:

Виктория Земляника лесная
Семейство Розоцветные:


Кустарниковая роза Чайная роза
Семейство Крыжовниковые:


Черная смородина Красная смородина
Семейство Лилейные:


Оранжевая лилия Белая садовая лилия
-
Полиморфизм популяций.
-
Изменение рисунка белых пятен на листьях клевера ползучего в зависимости от генотипа.
Изменения, происходящие в популяциях живых организмов под влиянием антропогенных факторов, используются для биоиндикации и мониторинга экосистем. Оценить состояние почвенного покрова и уровень антропогенного воздействия можно с помощью фенотипических биоиндикаторов. Степень реализации фенотипического разнообразия служит индикатором уровня благоприятствования условий среды.
Клевер ползучий (Trifolium repens L.) - многолетнее растение со стержневой, сильно разветвленной корневой системой, укороченным главным стеблем и боковыми стелющимися и укореняющимися побегами. Он хорошо растет на разных почвах с достаточным количеством питательных веществ и воды, но плохо переносит избыток влаги в почве, сухие и сильно-кислые почвы. Это светолюбивое растение, которое при благоприятных условиях быстро разрастается и
образует сомкнутый покров, вытесняя из травостоя злаки и разнотравье.
Характерной экологической особенностью клевера ползучего является хорошая адаптация к большому диапазону абиотических условий, в том числе и экстремальных, что способствует его широкому распространению на нарушенных территориях. Однако клевер не выдерживает конкуренции с другими видами и сохраняется в сообществах, находящихся под постоянной антропогенной нагрузкой. Он относительно устойчив к механическим повреждениям, прежде всего к вытаптыванию. Для клевера ползучего в естественных местах обитания лучше абиотические
условия, но острее ценотический стресс; в загрязненных местообитаниях, хотя и имеют место токсические эффекты нефтепродуктов, но практически полностью снята конкуренция.
Для клевера ползучего характерна вегетативно-подвижная жизненная форма, представленная особями семенного происхождения (генетами) и потомками вегетативного размножения разной степени разветвленности. Первые (генеты) встречаются почти исключительно на участках с нарушенным растительным покровом. В длительно существующих ненарушенных ценозах клевер представлен удлиненными плагиотропными побегами, нарастающими за счет верхушечной меристемы и активно образующими пазушные боковые побеги. Вегетативное размножение
способствует удержанию и распространению по территории, а генеративное за счет мощного банка семян служит захвату новых территорий при нарушении там растительного покрова.
Характерной особенностью природных популяций клевера ползучего является полиморфизм по форме седого рисунка (пятна) на листовой пластинке. Рисунок на листе может отличаться расположением,окраской, интенсивностью проявления, размером. На его выраженность оказывают влияние возраст,форма, относительный размер листьев. Доказано, что разнообразие растений по этому признаку определяется серией множественных аллелей гена V .
Наличие «седого» пятна на листьях - признак доминантный (V), его отсутствие - рецессивный (v).
Все аллели гена V нарушают нормальное развитие хлорофилла в палисадных клетках светлой зоны листа,приводят к сокращению в них количества хлоропластов вплоть до их полного отсутствия, способствуют уменьшению размеров палисадных клеток и увеличению пространства между ними, более ранней гибели клеток.
Различные уровни расположения пятен соответствуют времени действия соответствующих аллелей,нарушающих нормальное развитие хлорофилла, в онтогенезе. Насчитывается 11 или более аллелей этого гена. Для большинства комбинаций аллелей характерно их проявление совместно с образованием различных вариантов. Форма седого рисунка на пластинках листа клевера ползучего и частота его встречаемости - индикатор загрязнения среды обитания.
Изучению природных популяций клевера ползучего посвящен ряд исследований, в которых приводится анализ пространственной и возрастной структуры популяций, характеризуется эколого-генетическая и межпопуляционная изменчивость по признаку пятна на листьях.

Р.п. Карабаевка, Тагайский лес.
А) Закладка 20 площадок в тени и на солнце размером 25*25 см.


Площадка ( освещенное место) Площадка ( затененное место)
Б) Таблица изменения рисунка белых пятен у клевера:
 Генотип
vv ( освещенное место)
Генотип
vv ( освещенное место)
 Генотип
VV ( освещенное место)
Генотип
VV ( освещенное место)
 Генотип VHVH ( освещенное место)
Генотип VHVH ( освещенное место)
 Генотип
VHVH (затененное место)
Генотип
VHVH (затененное место)
 Генотип
vv( затененное место)
Генотип
vv( затененное место)
 Генотип
VV (затененное место)
Генотип
VV (затененное место)
Выводы:
-
На площадках в освещенном месте преобладает клевер, с белыми пятна с генотипом VHVH, на втором месте клевер с белыми пятнами с генотипом VV, а очень редко встречается клевер без белых пятен с генотипом vv.
-
На площадках в затененном участке преобладает клевер без белых пятен с генотипом vv, на втором месте клевер с белыми пятнами с генотипом VV, редко встречается клевер с белыми пятнами с генотипом VHVH.
-
Такие результаты связаны с тем, что белое пятно может быть результатом различной скорости роста эпидермальных клеток по сравнению с палисадными, вследствие чего палисадные клетки подвергаются более сильному воздействию света, с последующим разрушением хлорофилла.
-
Также белые пятна на клевера зависят и от антропогенного воздействия. Чем меньше воздействие, тем меньше вероятность проявления белых пятен.